

Application of 16SrRNA Gene Detection in the Early Diagnosis of Bacterial Meningitis in Children
-
摘要:
目的 探讨脑脊液(cerebrospinal fluid,CSF)中16SrRNA基因检测在儿童细菌性脑膜炎(BM)早期诊断中的应用价值。 方法 收集40例2019年1月至2020年6月间,在昆明市儿童医院确诊为BM患者的CSF标本进行16SrRNA基因PCR检测,16SrRNA基因PCR检测阳性标本进行基因测序,测序结果通过NCBI BLAST进行序列比对和同源性分析,同时进行CSF细菌培养,将CSF 16SrRNA基因PCR检测结果与CSF培养结果进行比较。 结果 40例患儿CSF中16例16SrRNA基因PCR检测阳性,阳性率40%;7例细菌培养阳性,阳性率17.5%,PCR检测法阳性率高于细菌培养法(χ2 = 4.93,P < 0.05),以CSF培养为“金标准”,PCR检测法的灵敏度为71.4%(5/7),特异度为66.7%(22/33);16SrRNA基因测序结果与CSF培养结果一致,且检测出CSF培养未检出的5种细菌;CSF培养时间为(61.21±12.62)h,PCR检测法所需时间为(7.09±0.45)h,两者之间差异有统计学意义(P < 0.05)。 结论 CSF中16SrRNA基因PCR检测法能提高BM患者CSF中病原菌的检出率,且能降低漏检率,具有特异、快速特点能及时为临床BM早期诊断提供可靠的病原学依据。 Abstract:Objective To explore the value of 16SrRNA gene detection in cerebrospinal fluid (CSF) in the early diagnosis of bacterial meningitis (BM) in children. Methods CSF specimens were collected from 40 patients with BM who were diagnosed of BM in Kunming Children’ s Hospital between January 2019 and June 2020. PCR was used to detect the 16SrRNA gene in the specimens. Then the 16SrRNA gene was sequenced for the positive samples by PCR, and the sequencing results sequence alignment and homology were analyzed by NCBI BLAST. At the same time, all the specimens were cultured simultaneously. Results Of the 40 children’ s CSF samples, 16 were positive for 16SrRNA gene PCR, with a positive rate of 40%; 7 cases were positive for bacterial culture, with a positive rate of 17.5%. The positive rate of PCR method was higher than that of the bacterial culture method (χ2 = 4.93, P < 0.05), with CSF as the “gold standard”, the sensitivity and specificity of PCR assay were 71.4% (5/7) and 66.7% (22/33); the results of 16SrRNA gene sequencing were consistent with the results of CSF culture, and five bacterias which were not detected by CSF culture; the time of CSF culture was (61.21±12.62) h, however, the PCR detection time required (7.09±0.45) h, and the difference of detection time between the two methods was statistically significant (P < 0.05). Conclusion The PCR detection method of 16SrRNA gene in CSF can improve the detection rate of CSF pathogenic bacteria in BM patients, and it can reduce the missed detection rate. It has the specific and rapid characteristics and can provide the reliable pathogenic basis for early diagnosis of clinical BM in time. -

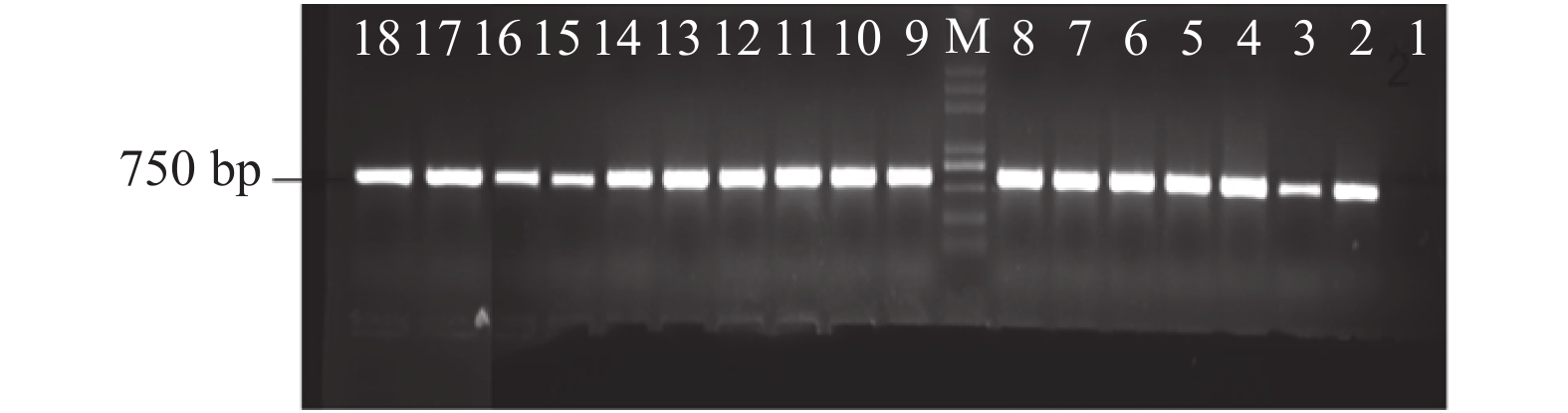
图 1 16例CSF临床样本16SrRNA基因PCR扩增产物琼脂糖凝胶电泳
M:DNA Marker;1:阴性对照(EBV);2:阳性对照(金黄色葡萄球菌);3~4:为大肠埃希菌;5:肺炎克雷伯杆菌;6:肺炎链球菌;7:无乳链球菌;3~7为两种方法检测均为阳性;8~18为细菌培养阴性,而16SrRNA基因PCR基因检测阳性标本。
Figure 1. Agarose gel electrophoresis of 16SrRNA gene PCR amplification products from clinical samples of 16 cerebrospinal fluid
表 1 40例细菌性脑膜炎患儿一般情况分析[n(%)]
Table 1. Analysis of general situations of 40 children with bacterial meningitis [n(%)]
项目 基本信息 总计 年龄 新生儿(< 28 d) 14(35.0) 28 d~1岁 16(40.0) 1~3岁 6(15.0) 大于3岁 4(10.0) 性别 男 17(42.5) 女 23(57.5) 临床表现 发热 30(75.0) 抽搐 6(15.0) 头痛 11(27.5) 呕吐 8(20.0) 腹泻 5(12.5) 病理征 4(10.0) 脑膜刺激征 7(17.5) 膝反射 4(10.0) 脑脊液改变 白细胞数(106/L) < 50 4(10.0) 50~500 13(32.5) 501~1000 5(12.5) > 1000 15(37.5) 腰穿前是否使用过抗生素 是 28(70) 否 12(30) 基础疾病 外伤 2(5.0) 手术后脑积水 5(12.5) 临床转归 康复或痊愈 29(72.5) 后遗症 5(12.5) 死亡/放弃治疗 1(2.5)  下载: 导出CSV
下载: 导出CSV
表 2 40例细菌性脑膜炎患儿血培养、CSF培养、CSF细菌涂片及CSF16SrRNA基因检测阳性率比较[n(%)]
Table 2. Comparison of blood culture,CSF culture,CSF bacterial smear and CSF 16SrRNA gene detection positive rate of 40 children with bacterial meningitis [n(%)]
方法 阳性率 χ2 P 血培养 6(15.0)* 脑脊液培养 7(17.5)* 脑脊液细菌涂片 4(10.0)* 12.94 0.005 脑脊液16SrRNA 基因PCR法 16(40.0) 与1616SrRNA基因检测法比较,*P < 0.05。
下载: 导出CSV
表 3 16SrRNA基因PCR检测结果与细菌培养结果比较(n)
Table 3. Comparison between 16SrRNA gene PCR and bacterial culture results (n)
16SrRNA基因PCR法 脑脊液细菌培养法 合计 χ2 P 阳性 阴性 阳性 5 11 16 阴性 2 22 24 4.93 0.026 合计 7 33 40
下载: 导出CSV
表 4 各年龄段两种不同方法检测病原菌分布情况(n)
Table 4. Two different methods to detect the distribution of pathogens in each age group (n)
病原菌 病原菌检测方法 < 28 d 28 d~1岁 1~3岁 ≥3岁 总计 脑脊液培养 16SrRNA基因PCR法 大肠埃希氏菌 4 6 3 3 0 0 6 肺炎克雷伯 1 1 0 0 1 0 1 肺炎链球菌 1 1 0 0 1 0 1 无乳链球菌 1 1 1 0 0 0 1 金黄色葡萄球菌 0 1 0 0 1 0 1 溶血性葡萄球菌 0 2 0 1 1 0 2 流感嗜血杆菌 0 2 1 0 1 0 2 表皮葡萄球菌 0 1 0 0 1 0 1 鞘氨醇单胞菌 0 1 0 0 0 1 1 总计 7 16 5 4 6 1 16
下载: 导出CSV
-
[1] A Sharew,J. Bodisen,B. R. Hansen,et al. The cause of death in bacterial meningitis[J]. BMC Infect Dis,2020,20(1):182-191. [2] Sanjith S,S M K,G T P,et al. Correlation between CSF glucose estimation using glucometers against the conventional laboratory technique in determining bacterial meningitis:An Indian study[J]. J Assoc Physicians India,2020,68(2):43-47. [3] 郑可鲁,杨思达,高媛媛,等. 脑脊液细菌16S核糖体RNA基因检测在抗生素治疗后化脓性脑膜炎患儿中的应用价值[J]. 广西医学,2016,38(10):1367-1369. [4] Ahmet S,Demet G Toprak S,et al. Evaluation of the line probe assay for therapid detection of bacterial meningitispathogens in cerebrospinal fluid samplesfrom children[J]. BMC Microbiology,2017,17(1):14-21. doi: 10.1186/s12866-016-0834-0 [5] 胡家胜,邓小龙,孙丹,等. 2016版《欧洲临床微生物和感染病学会急性细菌性脑膜炎诊治指南》解读[J]. 中国实用儿科杂志,2017,32(10):726-732. [6] 诸福棠. 实用儿科学[M]. 第8版. 北京: 人民卫生出版社, 2014: 854-1003. [7] 张勇,董红娟,梁静静,等. 成人急性细菌性脑膜炎临床特征和不良预后的危险因素分析[J]. 卒中与神经疾病,2020,27(4):509-512. doi: 10.3969/j.issn.1007-0478.2020.04.021 [8] 韩小妍,梁玉婵,郭晓珍,等. 病毒性脑炎和结核性脑膜炎的临床诊断鉴别分析[J]. 临床医学工程,2019,26(7):973-974. doi: 10.3969/j.issn.1674-4659.2019.07.0973 [9] 王海平,黄海林,麻明彪,等. 昆明地区新生儿化脓性脑膜炎分离菌分布及耐药性分析[J]. 世界最新医学信息文摘,2019,19(99):30-32,35. [10] 李阳,图雅,王育民. 儿童细菌性脑膜炎预后不良危险因素研究进展[J]. 中国实用神经疾病杂志,2020,23(15):1374-1380. [11] 吕其军,史太阳,周芳,等. 16S rRNA基因检测在脓毒症早期诊断中的研究进展[J]. 中华医院感染学杂志,2016,26(10):2395-2397. [12] 盛群英,林雅真,崔晓洁,等. 细菌16SrDNA测序分析妊娠晚期孕妇感染B族链球菌对阴道微生态及母婴结局的影响[J]. 国际检验医学杂志,2020,41(20):2528-2532. doi: 10.3969/j.issn.1673-4130.2020.20.024 [13] 陈文娟,干驰,赵瑞柯,等. PCR/16sRNA联合核苷酸测序法检测化脓性脑膜炎病原菌的临床价值[J]. 现代生物医学进展,2018,18(12):2289-2293. [14] 郑可鲁,杨思达,高媛媛,等. 抗生素治疗时限对化脓性脑膜患儿脑脊液细菌16srRNA基因检测的影响[J]. 临床与病理杂志,2016,36(9):1288-1292. [15] Vita S,Ajassa C,Caraffa E,et al. Immunological diagnosis as an adjunctive tool for an early diagnosis of tuberculous meningitis of an immune competent child in a low tuberculosis endemic country:acase report[J]. BMC Res Notes,2017,10(1):123-130. doi: 10.1186/s13104-017-2444-9 [16] Camara-Lemarroy C R,Delgado-Garcia G,et al. Mean platelet volumein the differential diagnosis of tuberculous and bacterial meningitis[J]. J Infect Dev Ctries,2017,11(2):166-172. doi: 10.3855/jidc.7478 -

 点击查看大图
点击查看大图
计量
- 文章访问数: 4501
- HTML全文浏览量: 2388
- PDF下载量: 10
- 被引次数: 0
