

Metformin Protects Type II Diabetic Kidneys by Regulating PARP-1 Activity
-
摘要:
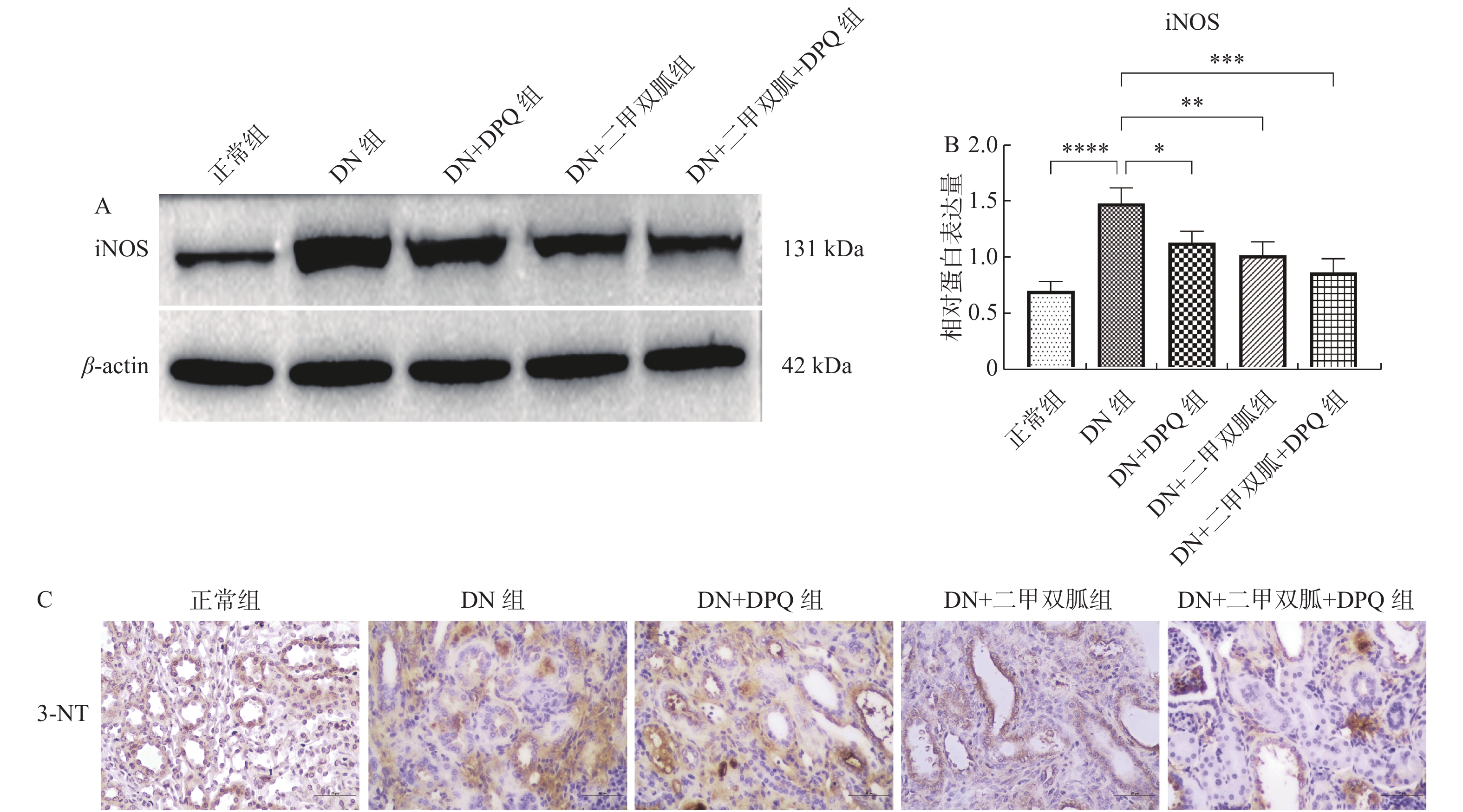
目的 研究二甲双胍通过调控多聚(ADP-核糖)聚合酶1(PARP-1)的活性,对2型糖尿病肾脏的保护作用及其机制。 方法 Wistar大鼠分为正常组(n = 12),DN组(n = 12),DN+DPQ组(n = 12),DN+二甲双胍组(n = 12)以及DN+二甲双胍+DPQ组(n = 12)。建模后检测各组大鼠空腹血糖含量,尿素氮含量,肌酐含量以及尿蛋白浓度等生化指标,HE染色和TUNEL染色观察肾脏病理情况,Western blot 检测PARP-1,iNOS,NF-κB及caspase-3的蛋白表达,ELISA检测炎性因子TNF-α及IL-1β的表达,免疫组化测定3-硝基酪氨酸(3-nitrotyrosine,3-NT)的表达。 结果 (1)3组治疗组大鼠的各项生化指标相较于DN组均有下降,其中DN + 二甲双胍 + DPQ组大鼠生化指标改变最为明显(P < 0. 01);(2)3组治疗组大鼠PARP-1的表达较于DN组下降,其中DN + 二甲双胍 + DPQ组的表达下降最为明显( P < 0.05);(3)3组治疗组大鼠肾脏组织的病理变化及肾脏细胞的凋亡较于DN组减缓,其中DN + 二甲双胍 + DPQ组的改善最为明显;(4)3组治疗组大鼠炎性因子的表达以及NF-kB,iNOS及3-NT的表达较于DN组下降,其中DN + 二甲双胍 + DPQ组的表达下降最为明显( P < 0.05)。 结论 二甲双胍通过调控PARP-1的表达,下调DN模型中NF-kB的表达,抑制NF-kB/iNOS/NO通路,抑制氧化损伤,减轻炎症反应,从而起到在糖尿病导致的高糖环境下,对肾脏的保护作用。 Abstract:Objective To investigate the protective effect of metformin on the kidney in type II diabetes mellitus (T2DM) by regulating the activity of poly (ADP-ribose) polymerase 1 (PARP-1) and investigate the mechanism of this effect. Methods Wistar rats were divided into normal group (n = 12), DN group (n = 12), DN + DPQ group (n = 12), DN + metformin group (n = 12) and DN + metformin + DPQ group (n = 12). After model establishment, biochemical parameters such as fasting glucose content, urea nitrogen content, creatinine content, and urine protein concentration were measured in each group of rats.HE staining and TUNEL staining were used to observe the renal pathology, Western blot was applied to detect the protein expression of PARP-1, iNOS, NF-κB and caspase-3.The expression of inflammatory factors TNF-α and IL-1β was detected by ELISA, and the expression of 3-nitrotyrosine (3-NT)was determined by immunohistochemistry. Results (1)The biochemical indexes of rats in the three treatment groups decreased compared with those in the DN group, with the most obvious changes in the DN + metformin + DPQ group (P < 0.01) ; (2) The expression of PARP-1 in rats in the three treatment groups decreased compared to the DN group, with the most significant decrease in the DN + metformin + DPQ group ( P < 0.05); (3) The pathological changes of kidney tissue and apoptosis of kidney cells in the three treatment groups were alleviatedcompared with the DN group, with the most significant decrease in the expression of DN + metformin + DPQ group; (4) The expression of inflammatory factors as well as the expression of NF-kB, iNOS and 3-NT decreased in the three treatment groups compared with the DN group, with the most significant decrease in the DN + metformin + DPQ group ( P < 0.05). Conclusion Metformin plays a role in protecting the kidney under the high glucose environment caused by diabetes through regulating the expression of PARP-1, down-regulating the expression of NF-kB in the DN model, inhibiting NF-kB/iNOS/NO pathway, inhibiting oxidative damage, and reducing inflammation. -
Key words:
- Diabetic nephropathy /
- Metformin /
- PARP-1 /
- Oxidative stress
-
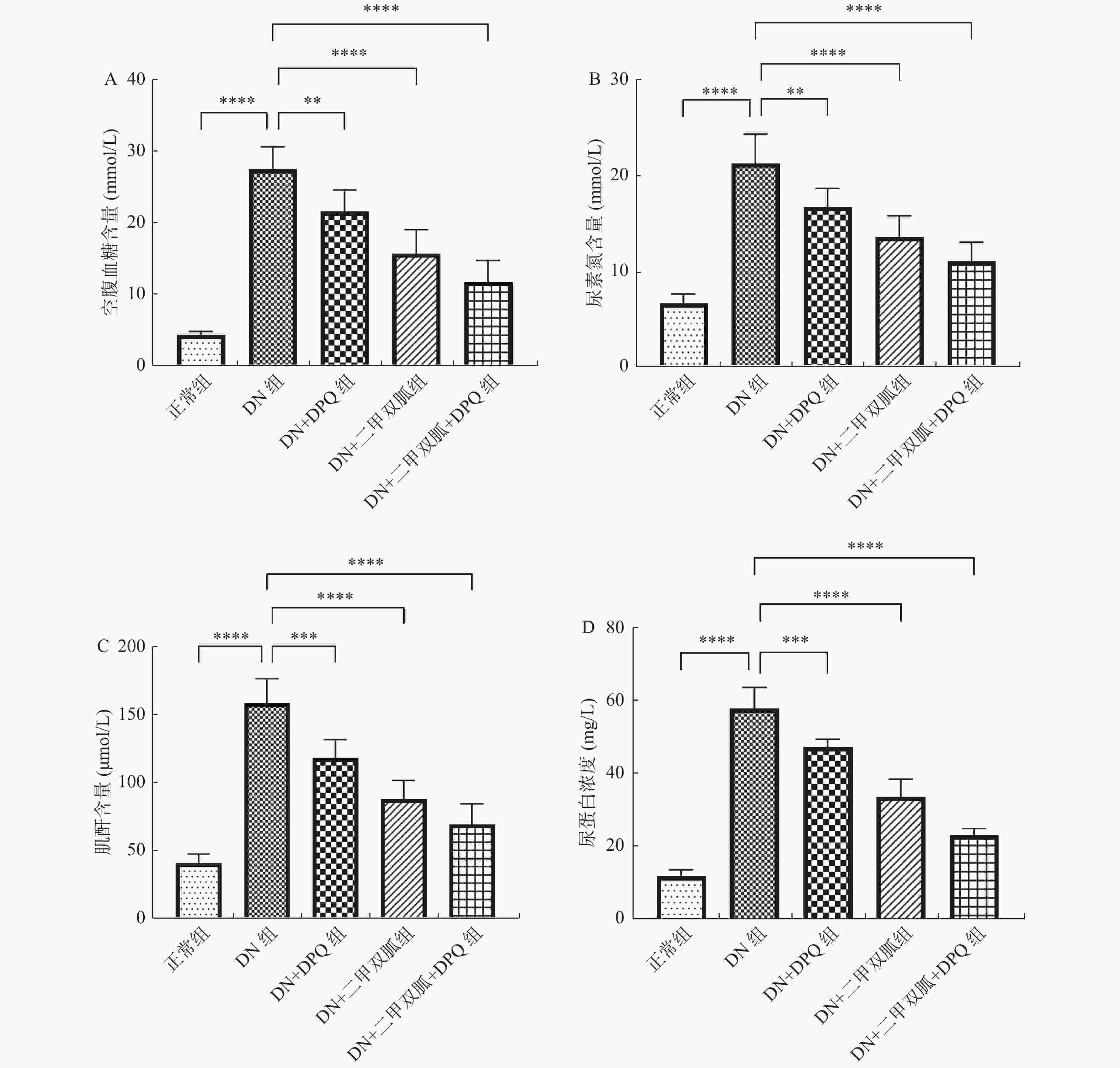
图 1 各组大鼠空腹血糖含量,尿素氮含量,肌酐含量以及尿蛋白浓度
A:空腹血糖含量;B:尿素氮含量;C:肌酐含量;D:尿蛋白浓度。两两组间比较,**P < 0.01,***P < 0.001,****P < 0.000 1。
Figure 1. Fasting glucose content,urea nitrogen content,creatinine content and urine protein concentration in each group of rats
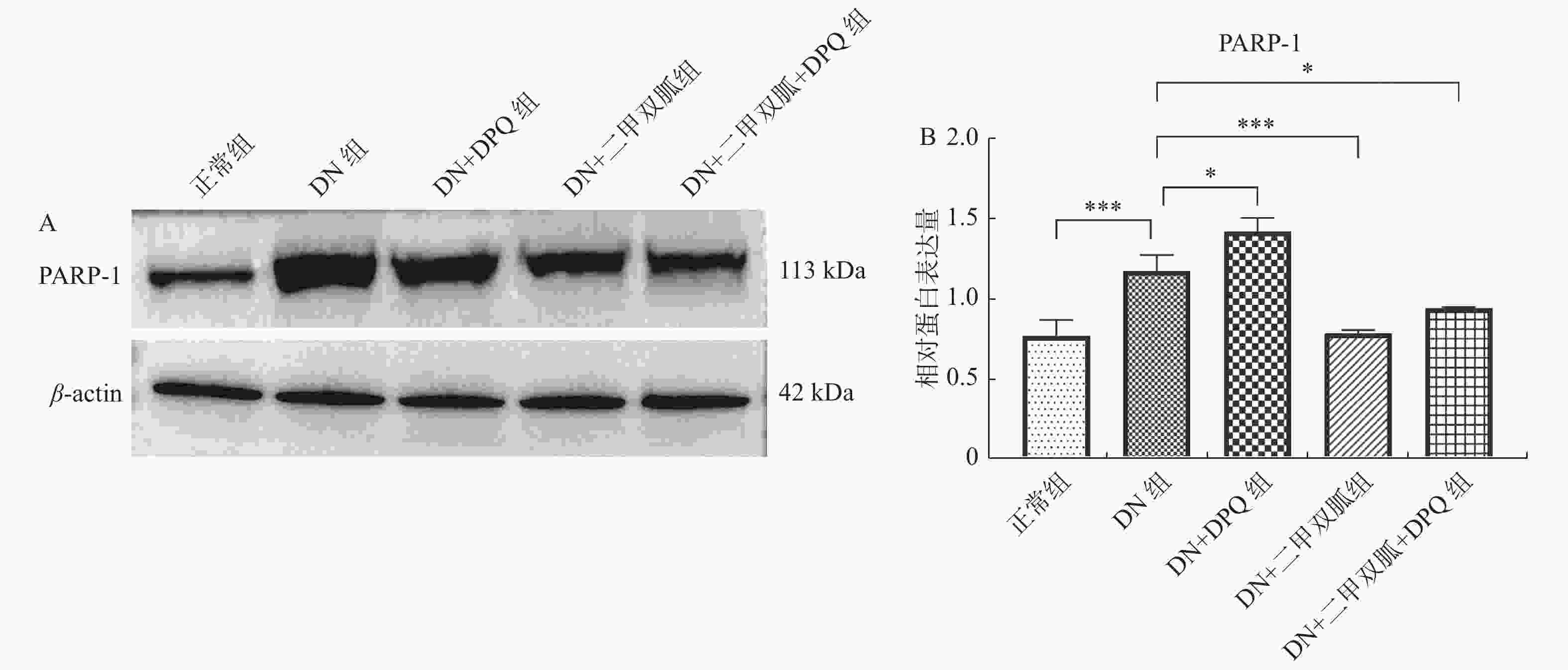
图 2 各组大鼠PARP-1相对蛋白表达
A:各组大鼠PARP-1蛋白表达条带;B:各组大鼠PARP-1相对蛋白表达量。两两组间比较,*P < 0.05,***P < 0.001,****P < 0.0001。
Figure 2. Relative expression of PARP-1 in each group
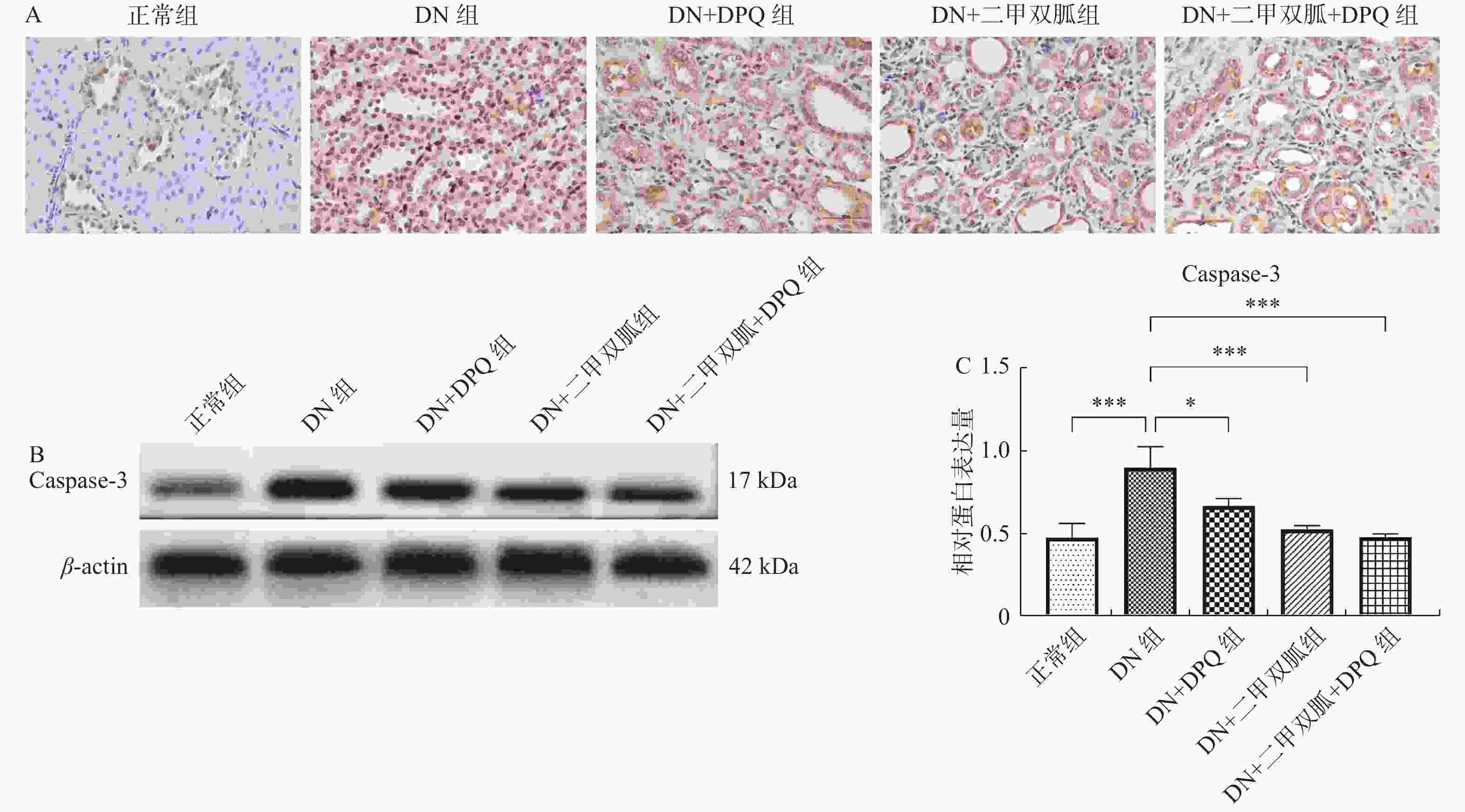
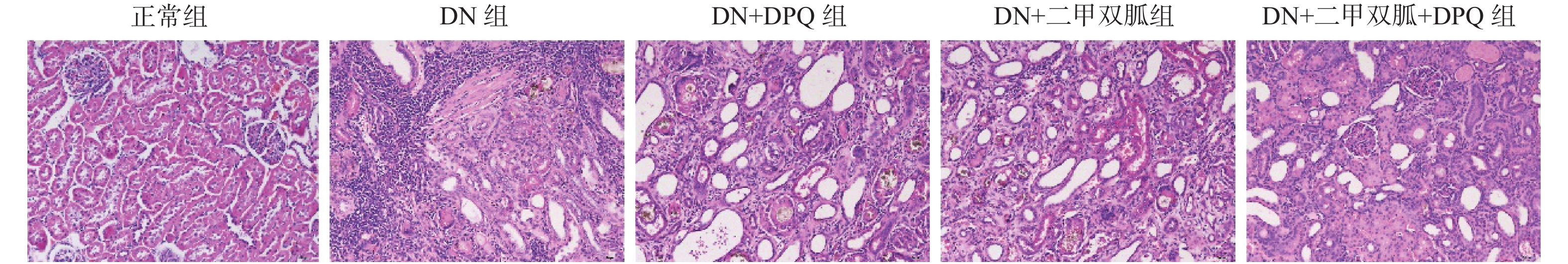
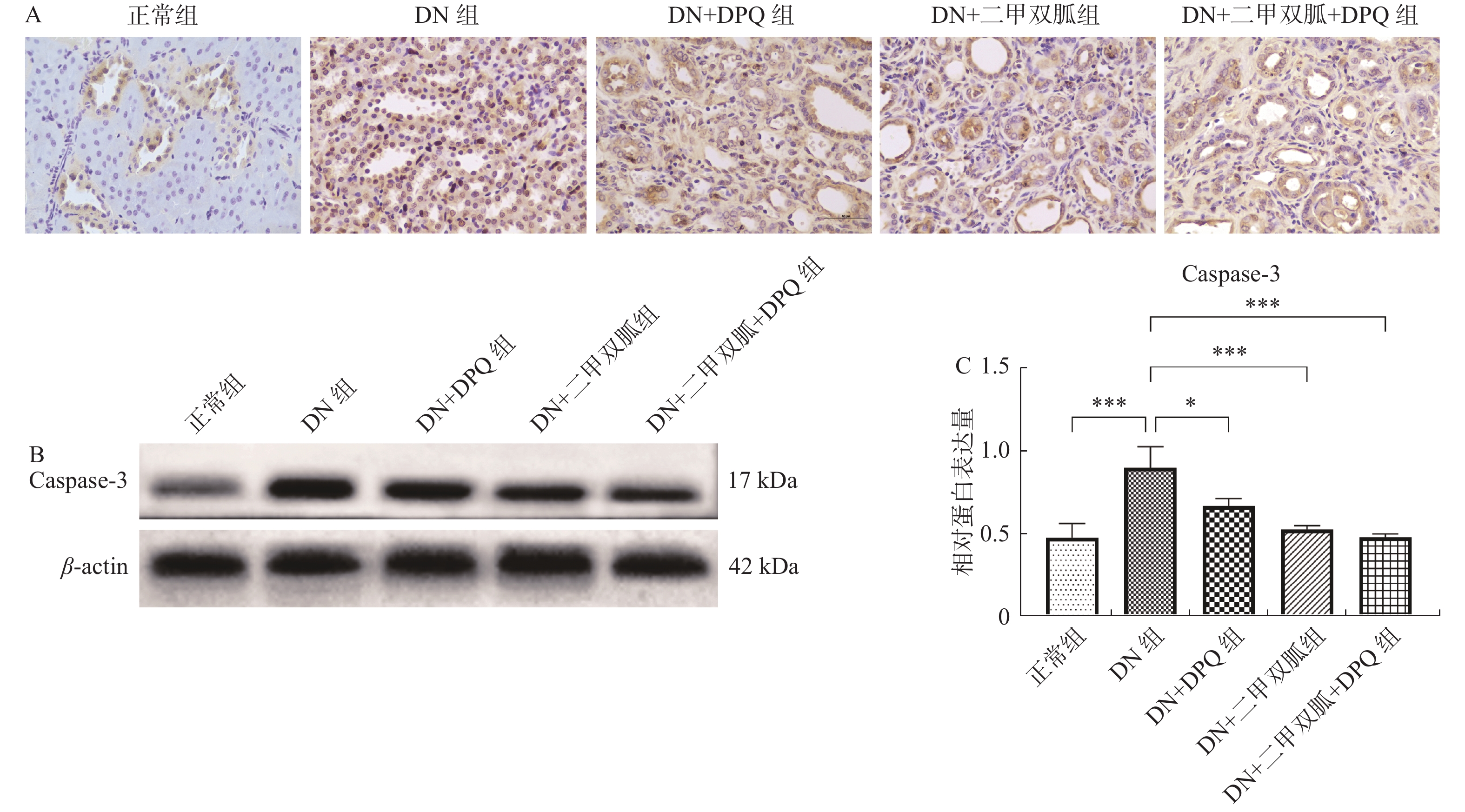
图 4 二甲双胍通过降低PARP-1表达减轻大鼠肾脏细胞凋亡
A:肾脏细胞凋亡(40*);B:Caspase-3 在各组中的蛋白表达条带;C:Caspase-3 在各组中的相对蛋白表达。两两组间比较,*P < 0.05,***P < 0.001,****P < 0.000 1。
Figure 4. Metformin attenuates renal cell apoptosis in rats by reducing PARP-1 expression
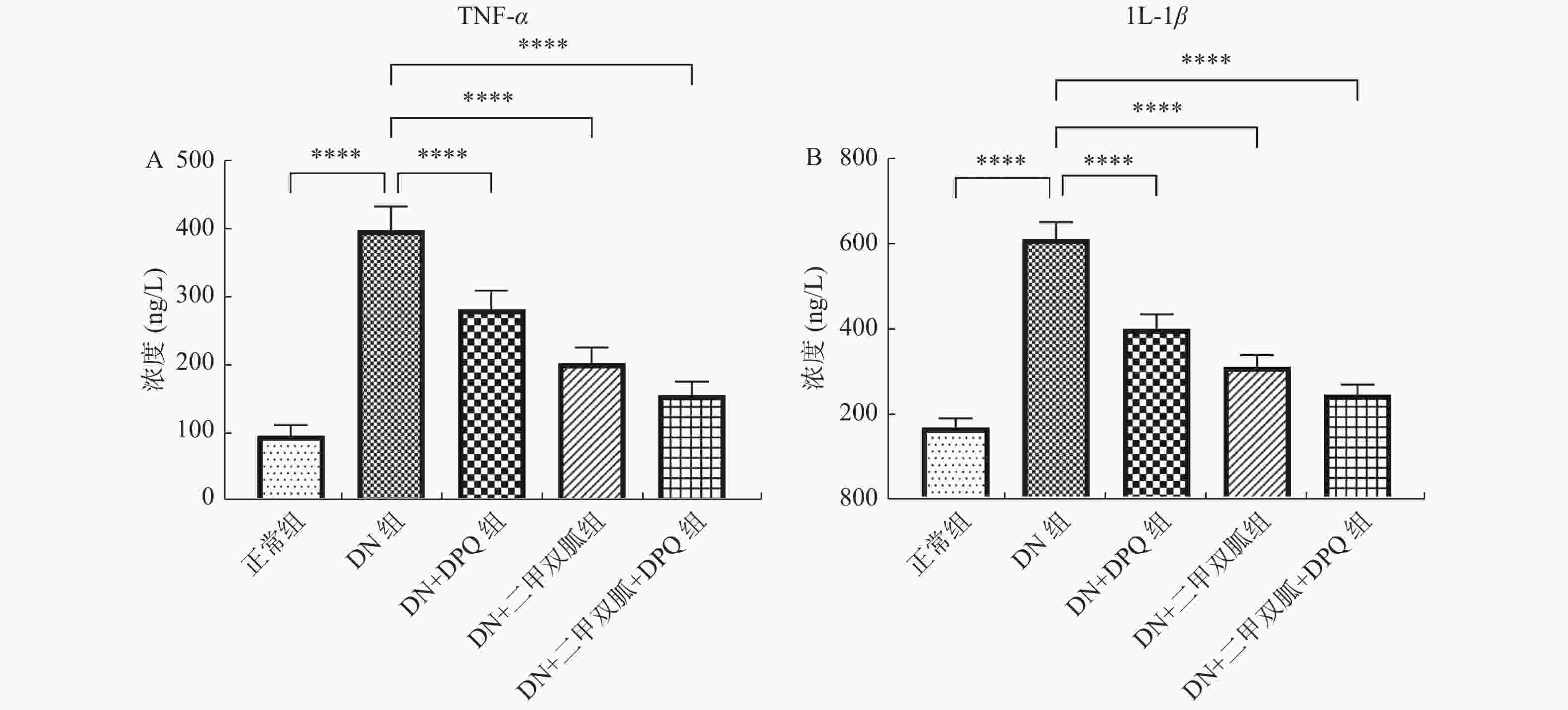
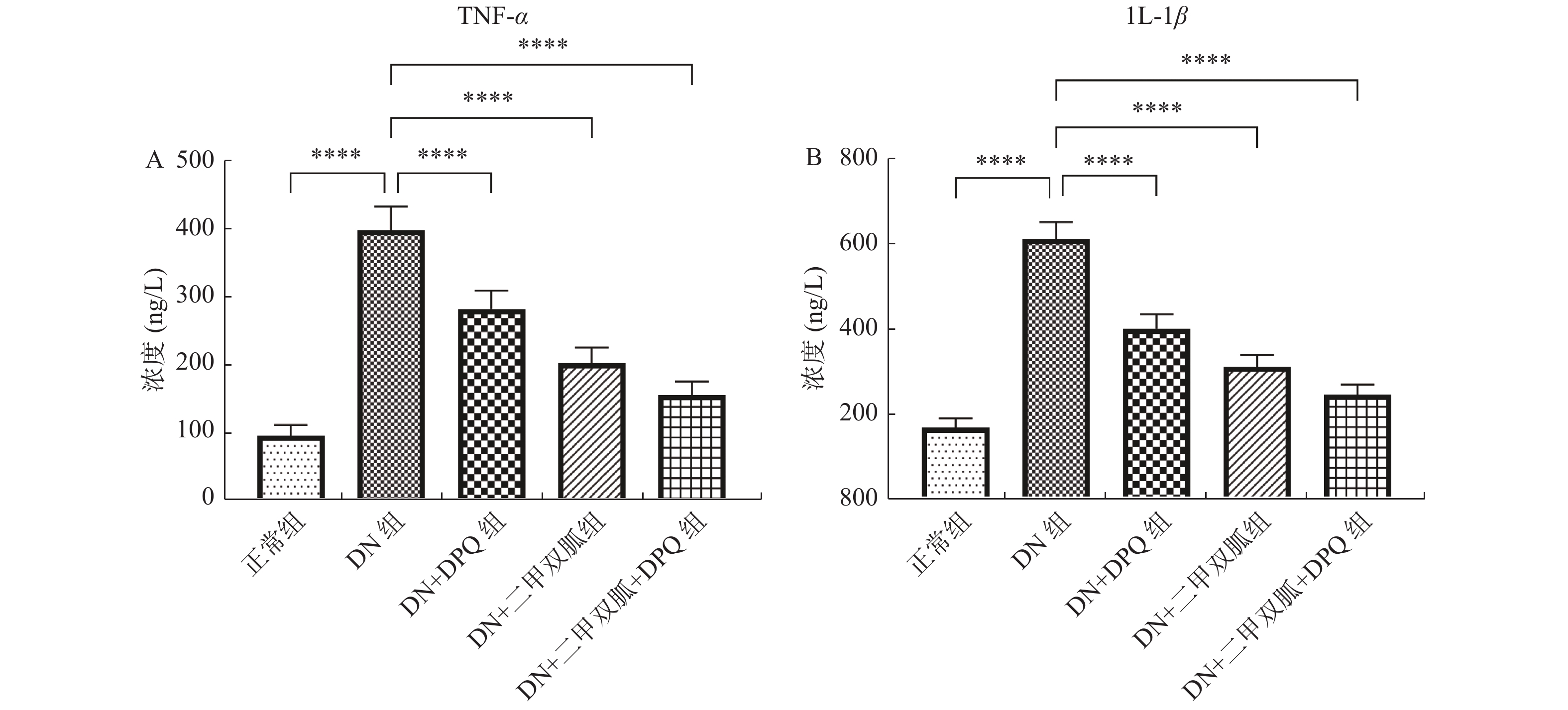
图 5 各组大鼠TNF-α及IL-1β浓度
A:各组大鼠TNF-α浓度;B:各组大鼠IL-1β浓度。两两组间比较,****P < 0.000 1。
Figure 5. Relative expression of TNF-α and IL-1β in each group
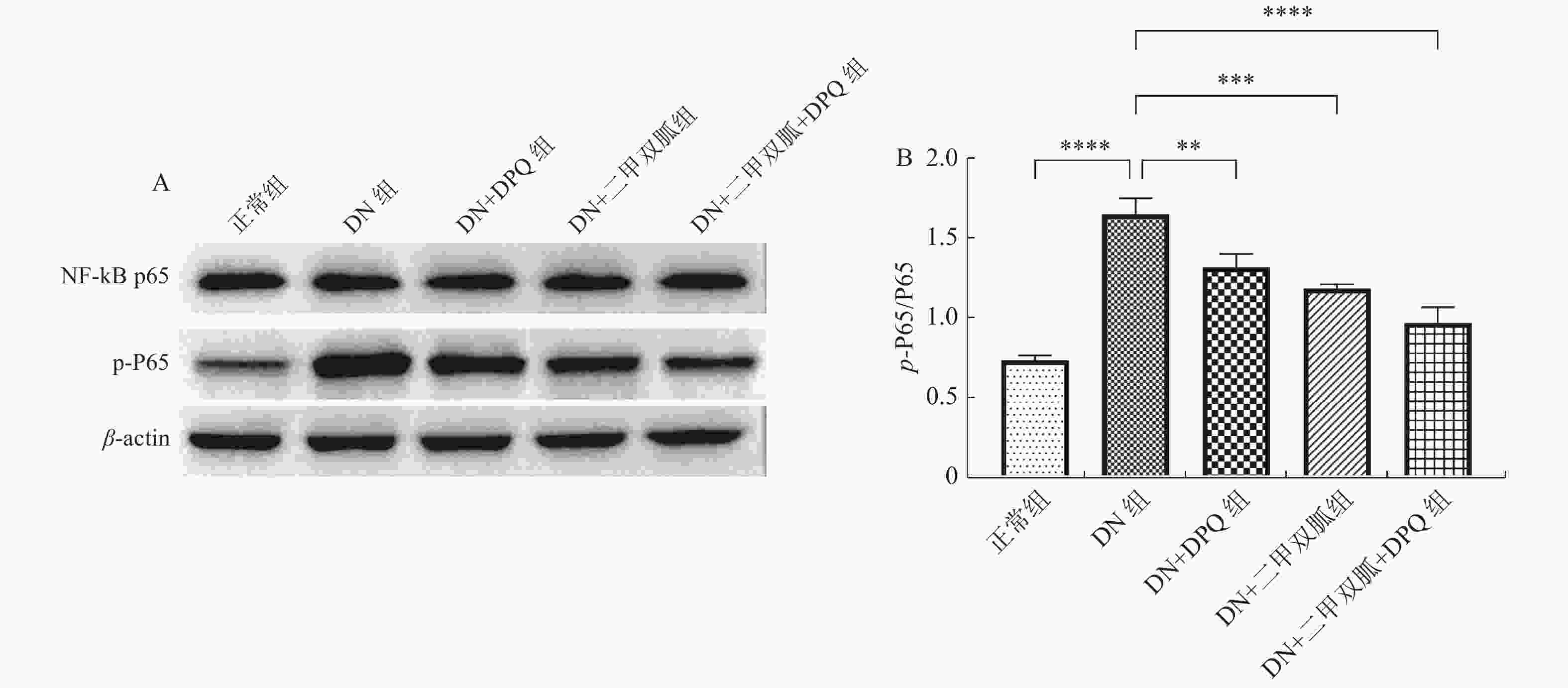
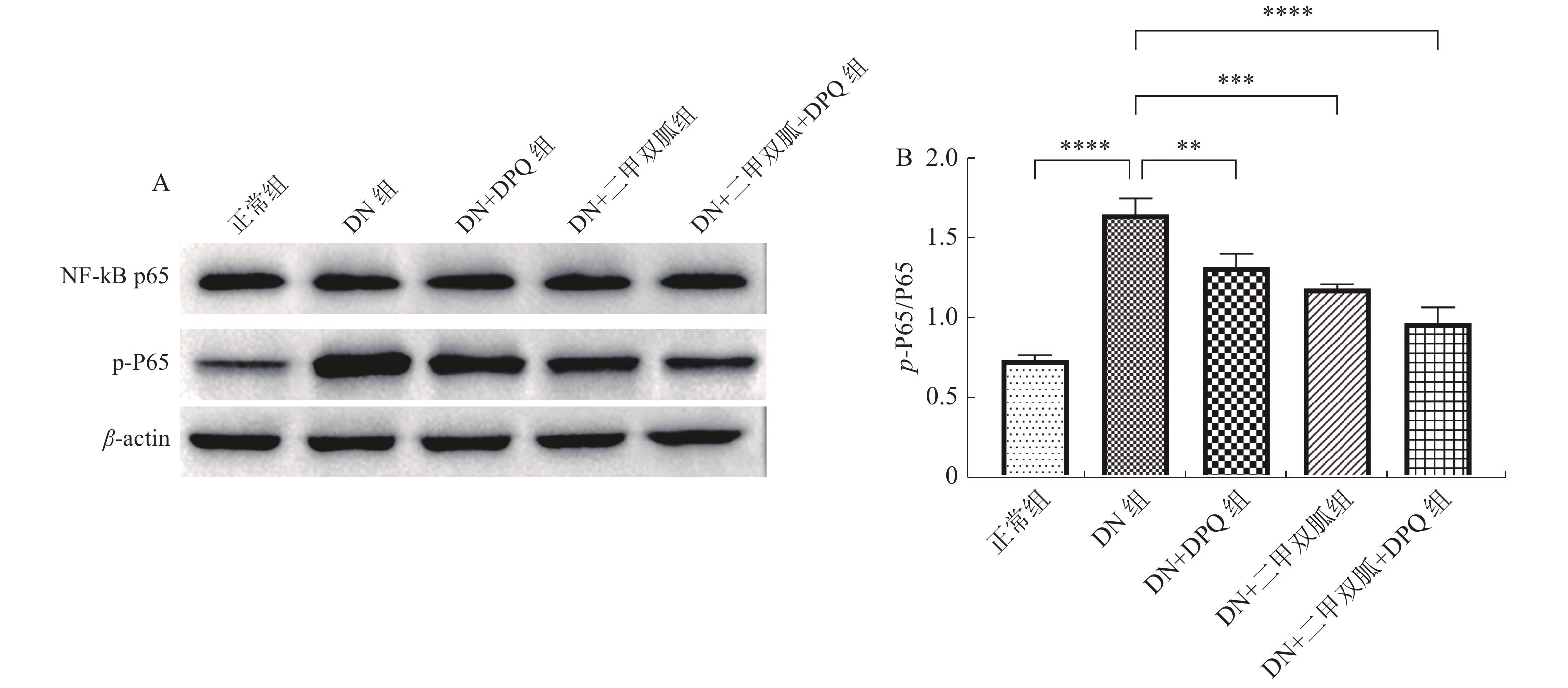
图 6 各组大鼠NF-κB相对蛋白表达
A:各组大鼠NF-κB p65 以及p-P65 蛋白表达条带;B:各组大鼠NF-κB p65 以及p-P65相对蛋白表达量。两两组间比较,**P < 0.01,***P < 0.001,****P < 0.000 1。
Figure 6. Relative expression of NF-κB in each group
-
[1] Li X,Li C,Li X,et al. Involvement of histone lysine methylation in p21 gene expression in rat kidney in vivo and rat mesangial cells in vitro under diabetic conditions[J]. J Diabetes Res,2016,2016(10):1-12. [2] 林善锬. 糖尿病肾病[J]. 中华内科杂志,2005,44(3):229-231. doi: 10.3760/j.issn:0578-1426.2005.03.028 [3] Rogacka D,Audzeyenka I,Rychowski M,et al. Metformin overcomes high glucose-induced insulin resistance of podocytes by pleiotropic effects on SIRT1 and AMPK[J]. Biochim Biophys Acta Mol Basis Dis,2017,1864(1):115-125. [4] Foretz M,Hébrard S,Leclerc J,et al. Metformin inhibits hepatic gluconeogenesis in mice independently of the LKB1/AMPK pathway via a decrease in hepatic energy state[J]. J Clin Invest,2010,120(7):2355-2369. doi: 10.1172/JCI40671 [5] El-Bassossy,Hany M. PARP-1 inhibition alleviates diabetic cardiac complications in experimental animals[J]. Eur J Pharmacol,2016,791:444-454. doi: 10.1016/j.ejphar.2016.09.008 [6] Guzyk M M,Tykhomyrov A A,Nedzvetsky V S,et al. Poly (ADP-Ribose) polymerase-1 (PARP-1) inhibitors reduce reactive gliosis and improve angiostatin levels in retina of diabetic rats[J]. Neurochem Res,2016,41(10):2526-2537. [7] Waldman M,Nudelman V,Shainberg A,et al. PARP-1 inhibition protects the diabetic heart through activation of SIRT1-PGC-1α axis[J]. Experimental Cell Research,2018,373(1-2):112-118. doi: 10.1016/j.yexcr.2018.10.003 [8] 陈小兰,颜安华,郑道国. 二甲双胍通过调控PARP-1活性对心肌梗死大鼠模型发挥保护作用[J]. 中国药师,2019,22(8):1401-1406. doi: 10.3969/j.issn.1008-049X.2019.08.006 [9] Anisimov V N J A. Metformin for aging and cancer prevention[J]. Aging (Albang NY),2010,2(11):760-774. [10] 管鑫娟,姚伟峰. 糖尿病肾病[J]. 解放军医学院学报,2012,17(1):116-117. [11] 杨倩,关天俊. 糖尿病肾病的诊治进展[J]. 世界临床药物,2018,39(2):93-97. [12] 孙志强. 糖尿病肾病研究进展[J]. 中华实用诊断与治疗杂志,2010,24(6):539-541. [13] 张舒媛,王东超,李博,等. 糖尿病肾病研究进展[J]. 世界中医药,2015(10):1621-1625. doi: 10.3969/j.issn.1673-7202.2015.10.044 [14] Denhez B,Rousseau M,Spino C,et al. Saturated fatty acids induce insulin resistance in podocytes through inhibition of IRS1 via activation of both IKKβ and mTORC1[J]. Sci Rep,2020,10(1):21628. doi: 10.1038/s41598-020-78376-1 [15] Su Y,Chen Q,Ju Y,et al. Palmitate induces human glomerular mesangial cells fibrosis through CD36-mediated transient receptor potential canonical channel 6/nuclear factor of activated T cell 2 activation[J]. Biochim Biophys Acta Mol Cell Biol Lipids,2020,1865(12):158793. doi: 10.1016/j.bbalip.2020.158793 [16] Xu Y,Gao H,Hu Y,et al. High glucose-induced apoptosis and necroptosis in podocytes is regulated by UCHL1 via RIPK1/RIPK3 pathway[J]. Exp Cell Res,2019,382(2):111463. doi: 10.1016/j.yexcr.2019.06.008 [17] Alicic R Z, Rooney M T, Tuttle K R. Diabetic kidney disease: Challenges, progress, and possibilities[J]. Clin J Am Soc Nephrol,2017,12(12):2032-2045. doi: 10.2215/CJN.11491116 [18] Lee M J,Feliers D,Mariappan M M,et al. A role for AMP-activated protein kinase in diabetes-induced renal hypertrophy[J]. Am J Physiol Renal Physiol,2007,292(2):F617. [19] Satriano J,Mansoury H,Deng A,et al. Transition of kidney tubule cells to a senescent phenotype in early experimental diabetes[J]. Am J Physiol Cell Physiol,2010,299(2):C374. [20] Shang F,Zhang J,Li Z,et al. Cardiovascular Protective Effect of Metformin and Telmisartan:Reduction of PARP1 Activity via the AMPK-PARP1 Cascade[J]. PLos One,2016,11(3):e0151845. [21] 耿彦平 王,谷剑. 二甲双胍对急性心肌梗死后心肌梗死面积及心功能的影响[J]. 临床荟萃,2016,31(7):805-808. [22] 林芳芳 张,牛姗姗. 二甲双胍预防糖尿病合并心肌梗死患者再梗死的效果观察[J]. 实用糖尿病杂志,2018,14(3):39. [23] 赵丽,梁晓春. 多聚(ADP-核糖)聚合酶与糖尿病周围神经病变[J]. 医学研究杂志,2011,40(8):23-25. doi: 10.3969/j.issn.1673-548X.2011.08.008 [24] 凌亦凌 朱 赵. 多聚(ADP-核糖)聚合酶与细胞损伤[J]. 中国病理生理杂志,2002,18(1):104-107. doi: 10.3321/j.issn:1000-4718.2002.01.033 [25] Pacher P,Szabo C. Role of the Peroxynitrite-Poly(ADP-Ribose)Polymerase Pathway in Human Disease[J]. Am J Palhol,2008,173(1):2-13. [26] Liang E S,Bai W W,Wang H,et al. PARP-1(Poly[ADP-Ribose] Polymerase 1)Inhibition Protects From Ang Ⅱ(Angiotensin Ⅱ)-Induced Abdominal Aortic Aneurysm in Mice[J]. Hypertension,2018,72(5):1189-1199. doi: 10.1161/HYPERTENSIONAHA.118.11184 [27] Chen H,Liu J,Wang B,et al. Protective effect of lncRNA CRNDE on myocardial cell apoptosis in heart failure by regulating HMGB1 cytoplasm translocation through PARP-1[J]. Arch Pharm Res,2020,43(12):1325-1334. doi: 10.1007/s12272-020-01290-7 [28] Luo H,Liang H,Chen Y,et al. miR-7-5p overexpression suppresses cell proliferation and promotes apoptosis through inhibiting the ability of DNA damage repair of PARP-1 and BRCA1 in TK6 cells exposed to hydroquinone[J]. Chem Biol Interact,2018,283:84-90. doi: 10.1016/j.cbi.2018.01.019 [29] Nan L,Xie Q,Chen Z,et al. Involvement of PARP-1/AIF Signaling Pathway in Protective Effects of Gualou Guizhi Decoction Against Ischemia-Reperfusion Injury-Induced Apoptosis[J]. Neurochem Res,2020,45(2):278-294. doi: 10.1007/s11064-019-02912-3 [30] Chowdhury P,Dey P,Ghosh S,et al. Reduction of metastatic potential by inhibiting EGFR/Akt/p38/ERK signaling pathway and epithelial-mesenchymal transition after carbon ion exposure is potentiated by PARP-1 inhibition in non-small-cell lung cancer[J]. BMC Cancer,2019,19(1):829. doi: 10.1186/s12885-019-6015-4 [31] Leng Y P,Qiu N,Fang W J,et al. Involvement of Increased Endogenous Asymmetric Dimethylarginine in the Hepatic Endoplasmic Reticulum Stress of Type 2 Diabetic Rats[J]. PLoS One,2014,9(2):e97125. [32] Lhotak S,Sood S,Brimble E,et al. ER stress contributes to renal proximal tubule injury by increasing SREBP-2-mediated lipid accumulation and apoptotic cell death[J]. Am J Physiol Renal Physiol,2012,303(2):F266. doi: 10.1152/ajprenal.00482.2011 [33] Lupachyk S,Watcho P,Stavniichuk R,et al. Endoplasmic Reticulum Stress Plays a Key Role in the Pathogenesis of Diabetic Peripheral Neuropathy[J]. Diabetes,2013,62(3):944-952. doi: 10.2337/db12-0716 [34] Paravicini T M,Touyz R M J D C. NADPH Oxidases,Reactive Oxygen Species,and Hypertension:Clinical implications and therapeutic possibilities[J]. Diabetes Care,2008,31(Supplement 2):S170-S180. [35] 李宏. PARP-1与氧化应激在糖尿病神经病变发病机制中的作用[J]. 检验医学与临床,2011,8(3):334-338. [36] 王燕萍,潘晓东,姜苏原,等. 氧化应激可通过NF-κB-iNOS-NO信号通路诱导小鼠胰岛β细胞凋亡[J]. 中国应用生理学杂志,2009,25(2):255-259. [37] 陆春丽,孙明谨,褚加成,等. 利拉鲁肽通过下调iNOS改善糖尿病前期小鼠胰岛功能[J]. 湖北医药学院学报,2018,37(4):17-20+105. [38] 苑天彤,冯丽辉,迟继铭,等. 何首乌提取物对糖尿病肾病早期大鼠肾组织iNOS和NT表达的影响[J]. 中国中医药科技,2015,22(3):280-283. -

 下载:
下载:

 点击查看大图
点击查看大图

计量
- 文章访问数: 4143
- HTML全文浏览量: 2272
- PDF下载量: 33
- 被引次数: 0
