

Effects of Silencing RND3 Expression on the Inflammatory Response and Apoptosis of Hippocampal Neurons Injured by Oxygen Glucose Deprivation/Reoxygenation
-
摘要:
目的 探讨沉默Rho家族GTP酶3(RND3)表达对氧糖缺失/复氧复糖(OGD/R)损伤海马神经细胞炎症反应和细胞凋亡的影响。 方法 使用包有沉默RND3表达的短发夹RNA(shRND3)及其阴性对照(shRND3-NC)序列的慢病毒载体分别对海马神经细胞进行转染。转染成功后对细胞进行2 h的缺氧缺糖培养再行复氧复糖建立OGD/R模型。CCK-8法检测细胞活力,RT-qPCR检测RND3及炎症和凋亡相关分子的mRNA表达,Elisa检测细胞上清液中炎症因子的浓度,Western Blot检测RND3和凋亡蛋白以及胞核内NF-κB的蛋白表达,Annexin V-FITC / PI染色和流式细胞仪检测细胞的凋亡率。 结果 与正常细胞相比,OGD/R损伤后细胞活力降低(P < 0.001),炎症因子(TNF-α、IL-6和IL-1β)和NF-κB表达升高(P < 0.001),促凋亡蛋白(Caspase-3和Bax)表达升高(P < 0.001),抗凋亡蛋白(Bcl-2)表达降低(P < 0.001),细胞的凋亡率升高(P < 0.001)。与沉默RND3阴性组相比,沉默RND3后,细胞活力增加(P < 0.01),炎症因子(TNF-α、IL-6和IL-1β)和NF-κB表达降低(P < 0.001),促凋亡蛋白(Caspase-3和Bax)表达降低(P < 0.001),抗凋亡蛋白(Bcl-2)表达升高(P < 0.001),细胞的凋亡率下降(P < 0.001)。 结论 沉默RND3表达通过抑制炎症反应和细胞凋亡减轻海马神经细胞的OGD/R损伤,沉默RND3抑制炎症反应的机制可能与抑制NF-κB通路有关。 -
关键词:
- Rho家族GTP酶3 /
- 氧糖缺失/复氧复糖 /
- 炎症反应 /
- 细胞凋亡 /
- NF-κB通路
Abstract:Objective To investigate the protective effect of silencing Rho family GTPase3 (RND3) expression against oxygen glucose deprivation/reoxygenation (OGD/R) injury in hippocampal neurons via inhibiting inflammation and apoptosis and its mechanism. Methods The OGD/R model was established after transfecting cells with lentiviral vectors containing the short hairpin RNA of silencing RND3 gene expression (shRND3) and its negative control (shRND3-NC) sequence. CCK-8 assay was used to detect cell viability. RT-qPCR was used to detect the gene expression of RND3 and inflammation-related and apoptotic molecules. ELISA was used to detect the protein concentration of inflammatory factors in the supernatant of cell culture. Western blot was used to detect the protein expression of RND3, apoptotic proteins and NF-κB of cell nucleus.Cell apoptosis rate was determined by Annexin V-FITC/PI staining and flow cytometry. Results Compared with normal cells, the cell viability was decreased after OGD/R injury, the expression of inflammatory factors and NF-κB was increased, the expression of pro-apoptotic proteins was increased, the expression of anti-apoptotic proteins was decreased, and the apoptosis rate of cells was increased. While compared with the negative RND3 silencing group, the cell viability was increased, the expression of inflammatory factors and NF-κB were decreased, the expression of pro-apoptotic protein was decreased, the expression of anti-apoptotic protein was increased, and the apoptosis rate of cells was decreased. Conclusion Silencing RND3 alleviates OGD/R injury in hippocampal neurons via inhibiting inflammatory response and apoptosis and the mechanism of inhibiting the inflammatory response by silencing RND3 may be related to the inhibition of NF-κB pathway. -

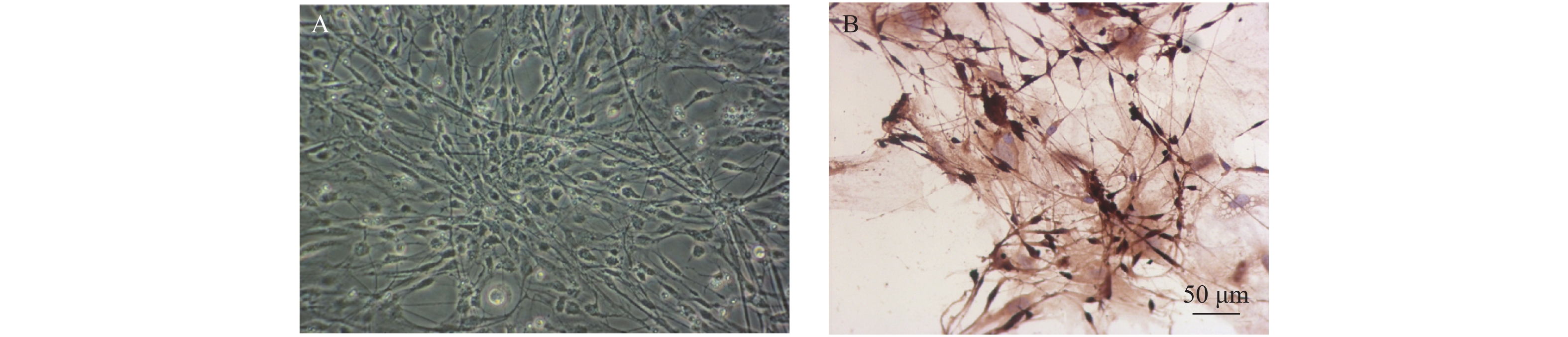
图 1 海马神经细胞的形态与鉴定
A:海马神经细胞典型形态(×200);B:海马神经细胞NeuN染色(比例尺:50 μm)。
Figure 1. Morphology and identification of hippocampal neurons

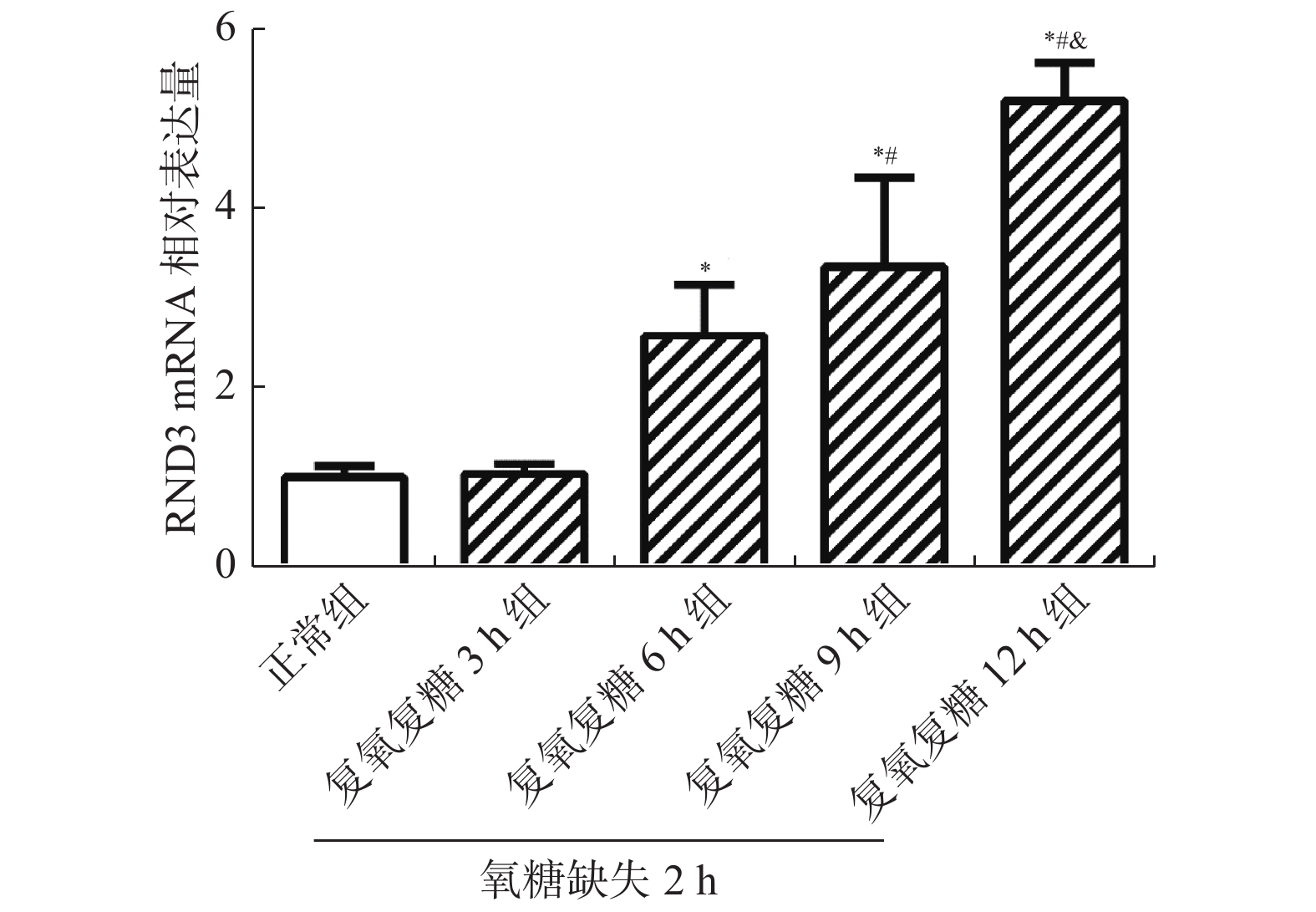
图 2 OGD/R后不同时刻海马神经细胞中RND3 mRNA的表达量
与正常组相比,*P < 0.05;与复氧复糖6 h组相比,#P < 0.05;与复氧复糖9 h组相比,&P < 0.05。
Figure 2. mRNA expression of RND3 in hippocampal neurons at different times after OGD/R

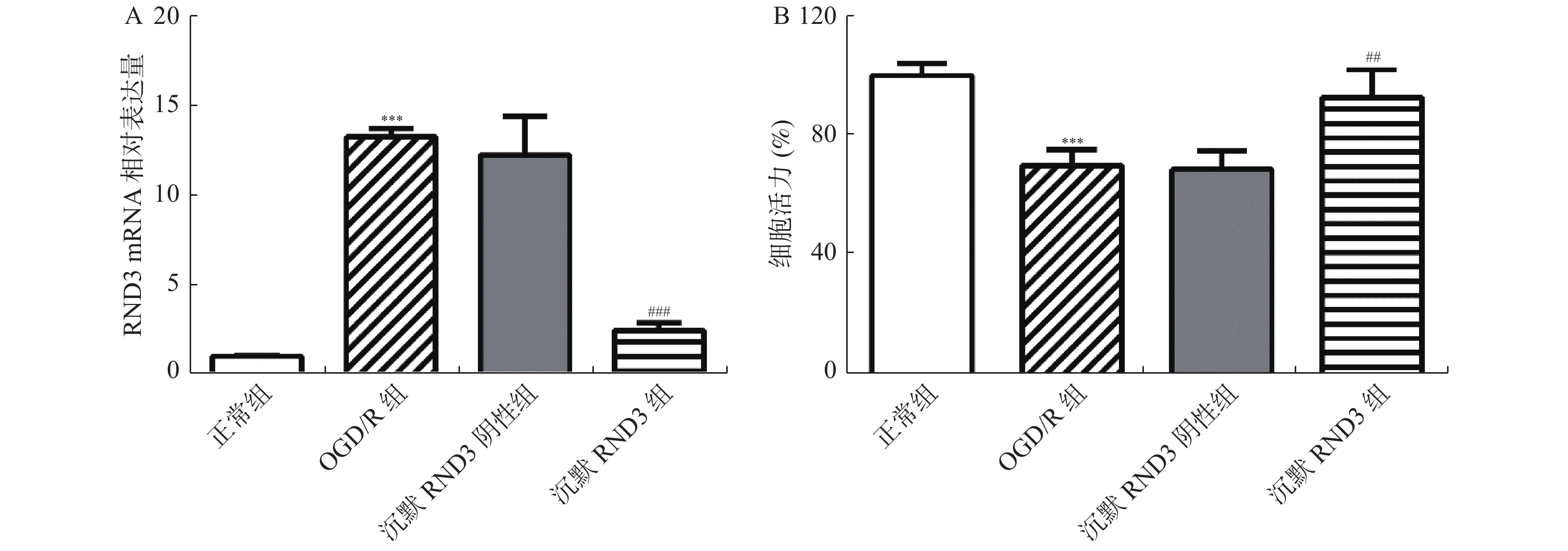
图 3 沉默RND3对OGD/R损伤海马神经细胞RND3 mRNA表达和细胞活力的影响
A:各组细胞RND的mRNA表达量;B:各组细胞的细胞活力;与正常组相比,***P < 0.001;与沉默RND3阴性组相比,##P < 0.01,###P < 0.001。
Figure 3. Effects of RND3 silencing on RND3 mRNA expression and cell viability of OGD/ R-injured hippocampal neurons

图 4 沉默RND3对OGD/R损伤海马神经细胞炎症因子表达的影响
A:各组细胞炎症因子TNF-α、IL-1β和IL-6的mRNA表达量;B:各组细胞上清液中炎症因子TNF-α、IL-1β和IL-6的蛋白浓度;与正常组相比,***P < 0.001;与沉默RND3阴性组相比,###P < 0.001。
Figure 4. The effect of silencing RND3 on the expression of inflammatory cytokines in OGD/R injured hippocampal neurons

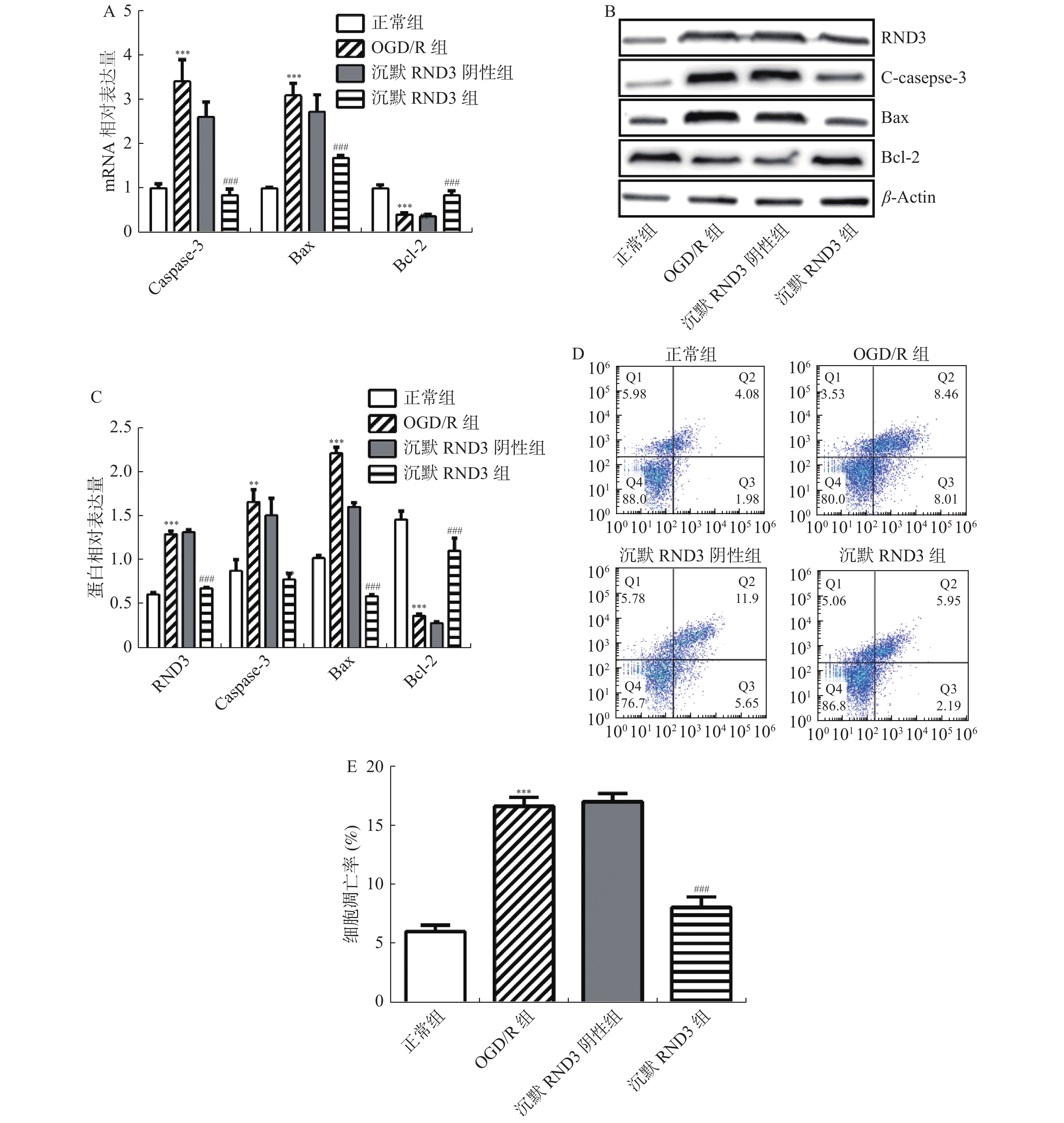
图 5 沉默RND3对OGD/R损伤海马神经元RND3蛋白表达和细胞凋亡的影响
A:各组细胞促凋亡蛋白Caspase-3、Bax和抗凋亡蛋白Bcl-2的mRNA相对表达量;B:各组细胞RND3蛋白,促凋亡蛋白C-caspase-3、Bax和抗凋亡蛋白Bcl-2的蛋白表达条带;C:各组细胞RND3蛋白,促凋亡蛋白C-caspase-3、Bax和抗凋亡蛋白Bcl-2的蛋白相对表达量;D:Annexin V-FITC / PI+流式细胞仪检测细胞凋亡;E:各组细胞的细胞凋亡率;与正常组相比,**P < 0.01,***P < 0.001;与沉默RND3阴性组相比,##P < 0.01,###P < 0.001。
Figure 5. Effects of silencing RND3 on RND3 protein expression and apoptosis in OGD/ R-injured hippocampal neurons

图 6 沉默RND3对OGD/R损伤海马神经元NF-κB(p65)表达的影响
A:各组细胞NF-κB(p65)mRNA表达量;B:各组细胞胞核内NF-κB(p65)的蛋白表达量;与正常组比较,***P < 0.001;与沉默RND3阴性组比较,###P < 0.001。
Figure 6. Effect of silencing RND3 on expression of NF-κB(p65)in OGD/ R-injured hippocampal neurons
表 1 引物序列
Table 1. The primer sequences
基因名称 上游引物序列(5′-3′) 下游引物序列(5′-3′) RND3 GCAAGATAGTAGTGGTGGGCG TCTGGGTAAGAGAGGGGACGG TNF-α CTCCTCACCCACACCGTCAG GAGCAGGTCCCCCTTCTCCA IL-1β TACAGTGGCAATGAGGATGAC CAAAGATGAAGGGAAAGAAGG IL-6 TTCGGTCCAGTTGCCTTCTC GTGCCTCTTTGCTGCTTTCA Caspase-3 CGGGCAAGCCAGATGTTTAT CAGCTCCGACTCTCCGAGAA Bax CCTTTTTGCTACAGGGTTTC TGTTGTTGTCCAGTTCATCG Bcl-2 GAGGGGCTACGAGTGGGATAC TCAGGCTGGAAGGAGAAGATG GAPDH CAAGTTCAACGGCACAGTCAAGG ACATACTCAGCACCAGCATCACC  下载: 导出CSV
下载: 导出CSV
-
[1] Goebel U, Scheid S, Spassov S, et al. Argon reduces microglial activation and inflammatory cytokine expression in retinal ischemia/reperfusion injury[J]. Neural Regen Res,2021,16(1):192-198. doi: 10.4103/1673-5374.290098 [2] Chai Z, Gong J, Zheng P, et al. Inhibition of miR-19a-3p decreases cerebral ischemia/reperfusion injury by targeting IGFBP3 in vivo and in vitro[J]. Biol Res,2020,53(1):17. doi: 10.1186/s40659-020-00280-9 [3] Tian Y, Zhong D, Liu Q, et al. Upregulation of miR-216a exerts neuroprotective effects against ischemic injury through negatively regulating JAK2/STAT3-involved apoptosis and inflammatory pathways[J]. J Neurosurg,2018,130(16):977-988. [4] Fang R, Zhao N, Zeng K, et al. MicroRNA-544 inhibits inflammatory response and cell apoptosis after cerebral ischemia reperfusion by targeting IRAK4[J]. Eur Rev Med Pharmacol Sci,2018,22(17):5605-5613. [5] Hao M, Xie L, Leng W, et al. Trim47 is a critical regulator of cerebral ischemia-reperfusion injury through regulating apoptosis and inflammation[J]. Biochem Biophys Res Commun,2019,515(4):651-657. doi: 10.1016/j.bbrc.2019.05.065 [6] Ballester-Lurbe B, Poch E, Mocholí E, et al. RhoE is spatiotemporally regulated in the postnatal mouse CNS[J]. Neuroscience,2009,163(2):586-593. doi: 10.1016/j.neuroscience.2009.06.062 [7] Jie W, Andrade K, Lin X, et al. Pathophysiological Functions of Rnd3/RhoE[J]. Compr Physiol,2015,6(1):169-186. [8] Guasch R, Blanco A, Pérez-Aragó A, et al. RhoE participates in the stimulation of the inflammatory response induced by ethanol in astrocytes[J]. Exp Cell Res,2007,313(17):3779-3788. doi: 10.1016/j.yexcr.2007.07.018 [9] Xu F, Lv C, Deng Y, et al. Icariside II, a PDE5 Inhibitor, suppresses oxygen-glucose deprivation/reperfusion-induced primary hippocampal neuronal death through activating the PKG/CREB/BDNF/TrkB signaling pathway[J]. Front Pharmacol,2020,11:523. doi: 10.3389/fphar.2020.00523 [10] Huang W, Liu X, Cao J, et al. miR-134 regulates ischemia/reperfusion injury-induced neuronal cell death by regulating CREB signaling[J]. J Mol Neurosci,2015,55(4):821-829. doi: 10.1007/s12031-014-0434-0 [11] Leng J, Liu W, Li L, et al. MicroRNA-429/Cxcl1 Axis Protective Against Oxygen Glucose Deprivation/Reoxygenation-Induced Injury in Brain Microvascular Endothelial Cells[J]. Dose Response,2020,18(2):710604679. [12] Zhang Z, Liu S, Huang S. Thymosin β4 prevents oxygen-glucose deprivation/reperfusion-induced injury in rat cortical neurons[J]. Neuropsychiatr Dis Treat,2019,15:2385-2393. doi: 10.2147/NDT.S208600 [13] Rao G, Zhang W, Song S. MicroRNA-217 inhibition relieves cerebral ischemia/reperfusion injury by targeting SIRT1[J]. Mol Med Rep,2019,20(2):1221-1229. [14] Liu A, Zhang W, Wang S, et al. HMGB-1/RAGE signaling inhibition by dioscin attenuates hippocampal neuron damage induced by oxygen-glucose deprivation/reperfusion[J]. Exp Ther Med,2020,20(6):231. [15] Yang Y, Hu F, Yang G, et al. Lack of sphingomyelin synthase 2 reduces cerebral ischemia/reperfusion injury by inhibiting microglial inflammation in mice[J]. Exp Ther Med,2020,20(6):241. [16] Li Y, Shen R, Wen G, et al. Effects of ketamine on levels of inflammatory cytokines IL-6, IL-1β, and TNF-α in the hippocampus of mice following acute or chronic administration[J]. Front Pharmacol,2017,8:139. [17] Zhou J, Zhou Q, Zhang T, et al. C7ORF41 Regulates Inflammation by Inhibiting NF-κB Signaling Pathway[J]. Biomed Res Int,2021,2021:7413605. [18] Moser B, Hochreiter B, Basílio J, et al. The inflammatory kinase IKKα phosphorylates and stabilizes c-Myc and enhances its activity[J]. Mol Cancer,2021,20(1):16. doi: 10.1186/s12943-021-01308-8 [19] Wang Q, Zhou X, Yang L, et al. Gentiopicroside (GENT) protects against sepsis induced by lipopolysaccharide (LPS) through the NF-κB signaling pathway[J]. Ann Transl Med,2019,7(23):731. doi: 10.21037/atm.2019.11.126 [20] Niu Y, Zhang W, Wu P, et al. Expression of the apoptosis-related proteins caspase-3 and NF-kappaB in the hippocampus of Tg2576 mice[J]. Neurosci Bull,2010,26(1):37-46. doi: 10.1007/s12264-010-6122-3 [21] Jiang D, Sun X, Wang S, et al. Upregulation of miR-874-3p decreases cerebral ischemia/reperfusion injury by directly targeting BMF and BCL2L13[J]. Biomed Pharmacother,2019,117:108941. doi: 10.1016/j.biopha.2019.108941 [22] Ma X, Li X, Tian F, et al. Upregulation of RND3 Affects Trophoblast Proliferation, Apoptosis, and Migration at the Maternal-Fetal Interface[J]. Front Cell Dev Biol,2020,8:153. doi: 10.3389/fcell.2020.00153 [23] Dong H, Sun Q, Zhang Y, et al. Genetic deletion of Rnd3 suppresses apoptosis through NF-κB signaling in the brain[J]. Oncol Rep,2021,45(2):595-605. -



 点击查看大图
点击查看大图
计量
- 文章访问数: 3712
- HTML全文浏览量: 2087
- PDF下载量: 24
- 被引次数: 0
