

Methamphetamine Alters Methylation Modifications of Synaptic Plasticity Genes in Addicted Mice
-
摘要:
目的 研究慢性甲基苯丙胺(methamphetamine,MA)成瘾小鼠神经突触可塑性基因的表达变化情况。 方法 选用C57BL/6J小鼠,模拟人类药物成瘾模式,分段渐进性腹腔注射给药5 mg/kg、10 mg/kg或者生理盐水。选取小鼠的大脑皮质和海马组织,通过亚硫酸氢盐处理基因组DNA和甲基化特异性PCR (methylmion specific PCR,MSP)对突触可塑性基因表达水平进行检测,运用BiQ-Analyzer软件对测序结果进行比对和DNA甲基化分析。 结果 相比盐水处理组小鼠,MA成瘾组小鼠大脑皮层的Egr2 (P = 0.064)基因启动子CpG位点的甲基化修饰增加,而Eln(P = 0.083)基因启动子的甲基化修饰降低;海马组织中,Arc(P = 0.025)、Egr2(P = 0.034)基因甲基化修饰增加,而Eln(P = 0.063)基因甲基化修饰降低。 结论 突触可塑性基因的甲基化修饰可能参与MA成瘾机制的形成。 Abstract:Objective To investigate the expression changes of synaptic plasticity genes in chronic methamphetamine (MA) addicted mice. Methods C57BL/6J mice were selected to simulate the human drug addiction model, and the drugs were injected intraperitoneally with 5 mg/kg, 10 mg/kg or normal saline. The cerebral cortex and hippocampus of mice were selected, and the gene expression level of synaptic plasticity was detected by bisulfite treated genomic DNA and methylation specific PCR (MethylmionSpecificPCR, MSP). The sequencing results were compared and DNA methylation was analyzed by BiQ-Analyzer software. Results Compared to mice in the saline-treated group, MA addiction group mice showed increased methylation modification of the Egr2 (P = 0.064) gene promoter CpG site and decreased methylation modification of the Eln (P = 0.083) gene promoter in the cerebral cortex; in hippocampal tissue, methylation modification of the Arc (P = 0.025) and Egr2 (P = 0.034) genes was increased, while Eln (P = 0.063) gene methylation modifications were decreased. Conclusion Methylation of synaptic plasticity genes may be involved in the formation of MA addiction mechanism. -
Key words:
- Methamphetamine /
- Synaptic plasticity /
- Immediate early gene /
- DNA Methylation.
-

图 1 生理盐水处理组和MA处理组大脑皮质和海马组织CpG岛甲基化修饰对比
A:生理盐水处理组和MA处理组大脑皮质基因组和海马组织基因组Arc启动子BSP测序结果,在海马组织中Arc(P < 0.05);B:生理盐水处理组和MA处理组大脑皮质基因组和海马组织基因组Fos启动子BSP测序结果;C:生理盐水处理组和MA处理组大脑皮质基因组和海马组织基因组Egr2启动子BSP测序结果,在海马组织中Egr2(P < 0.05);D:生理盐水处理组和MA处理组大脑皮质基因组和海马组织基因组Egr4启动子BSP测序结果。甲基化的CpG位点用实心圆圈表示、非甲基化的CpG位点用空心圆圈表示。一个测序克隆为一行。
Figure 1. Comparison of Methylation modification of CpG islands in cerebral cortex and hippocampus between saline treated group and MA treated group

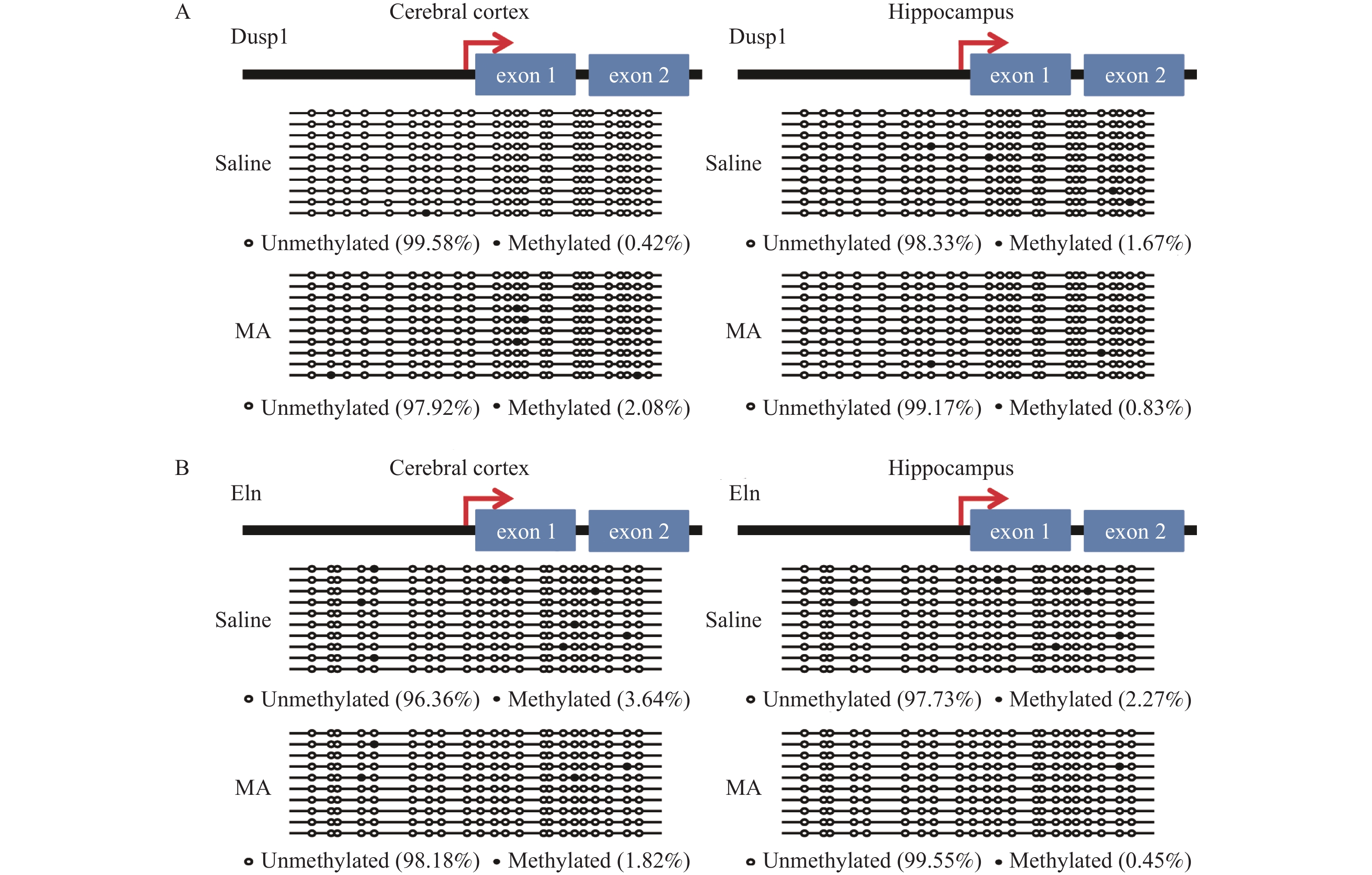
图 2 生理盐水处理组和MA处理组大脑皮质和海马组织CpG岛甲基化修饰对比
A:生理盐水处理组和MA处理组海马组织基因组Dusp1启动子BSP测序结果;B:生理盐水处理组和MA处理组海马组织基因组Eln启动子BSP测序结果,在海马组织中Eln(P = 0.063)。甲基化的CpG位点用实心圆圈表示、非甲基化的CpG位点用空心圆圈表示。一个测序克隆为一行。
Figure 2. Comparison of CpG island Methylation modifications in hippocampal tissues between the saline group and the MA treatment group
表 1 BSP引物表(5′→3′)
Table 1. Primer list for BSP (5′→3′)
基因 正向引物 反向引物 Fos GTGGGTAAGTTTTATTTTAGGA TAAAAAATCTCCTAAACCTTCC Arc GTATATTGTGTGGTTAGGATGG TAAAACCCTAACTCCCATTAAC Egr4 AATAAATTGGTATTAGGTGGGAAT CAATCCCCCCACTTATATAACTA Egr2 TTTAGTTTGGGTAGGGAAGAAA CTAACTTCCCTACTCCCCATT Dusp1 GGTGGGAATTAATTGGAAGTAT CACCCTCCTATCCTCTAAACTC Eln AGGAGGTTTGTAAGTTTGGTTTT AAATTCCCCATCAATCTTACC  下载: 导出CSV
下载: 导出CSV
-
[1] United Nations Office on Drugs and Crime. World drug report 2020[Z]. 2020. https://wdr.unodc.org/2020/. [2] 周凯程,何腾,刘馨雅,等. 甲基苯丙胺对小脑神经毒性的作用机制[J]. 中国药物依赖性杂志,2020,29(5):335-337,370. [3] Cheng M C,Hsu S H,Chen C H. Chronic methamphetamine treatment reduces the expression of synaptic plasticity genes and changes their DNA methylation status in the mouse brain[J]. Brain Research,2015,1629(10):126-134. [4] 王采玲. 甲基苯丙胺成瘾联合学习记忆细胞的发现及机制研究[D]. 南京: 南京中医药大学硕士论文, 2020. [5] Faure J J,Hattingh S M,Stein D J,et al. Proteomic analysis reveals differentially expressed proteins in the rat frontal cortex after methamphetamine treatment[J]. Metabolic Brain Disease,2009,24(4):685-700. doi: 10.1007/s11011-009-9167-0 [6] Ding X,Lee S W. Cocaine addiction related reproducible brain regions of abnormal default-mode network functional connectivity:A group ICA study with different model orders[J]. Neuroscience Letters,2013,5(29):110-114. [7] Day J J,Sweatt J D. DNA methylation and memory formation[J]. Nature Neuroscience,2010,13(11):1319-1323. doi: 10.1038/nn.2666 [8] Godino A,Jayanthi S,Cadet J L. Epigenetic landscape of amphetamine and methamphetamine addiction in rodents[J]. Epigenetics,2015,10(7):574-580. doi: 10.1080/15592294.2015.1055441 [9] Mccoy M T,Jayanthi S,Wulu J A,et al. Chronic methamphetamine exposure suppresses the striatal expression of members of multiple families of immediate early genes (IEGs) in the rat:Normalization by an acute methamphetamine injection[J]. Psychopharmacology,2011,215(2):353-365. doi: 10.1007/s00213-010-2146-7 [10] Jw A,Da A,Bs B,et al. Induction of immediate early genes expression in the mouse striatum following acute administration of synthetic cathinones[J]. Pharmacological Reports,2019,71(6):977-982. doi: 10.1016/j.pharep.2019.05.011 [11] Keiichiro M,Mika A,Hiroyuki O. Role of immediate-early genes in synaptic plasticity and neuronal ensembles underlying the memory trace[J]. Frontiers in Molecular Neuroscience,2016,8(8):78. [12] 郭敏,李刚. 突触可塑性相关蛋白的研究进展[J]. 神经药理学报,2013,3(6):57-64. [13] Kauer J A,Malenka R C. Synaptic plasticity and addiction[J]. Nature reviews Neuroscience,2007,8(11):844-858. doi: 10.1038/nrn2234 [14] L ü scher C,Malenka R C. Drug-evoked synaptic plasticity in addiction:From molecular changes to circuit remodeling[J]. Neuron,2011,69(4):650-663. doi: 10.1016/j.neuron.2011.01.017 [15] Zi ó Łkowska B,Gieryk A,Solecki W,et al. Temporal and anatomic patterns of immediate-early gene expression in the forebrain of C57BL/6 and DBA/2 mice after morphine administration[J]. Neuroscience,2015,284(9):107-124. [16] Thiriet N,Zwiller J,Ali S F. Induction of the immediate early genes egr-1 and c-fos by methamphetamine in mouse brain[J]. Brain Research,2001,919(1):31-40. doi: 10.1016/S0006-8993(01)02991-2 [17] Piechota M,Korostynski M,Solecki W,et al. The dissection of transcriptional modules regulated by various drugs of abuse in the mouse striatum[J]. Genome Biology,2010,11(5):48. doi: 10.1186/gb-2010-11-5-r48 [18] Robison A J,Nestler E J. Transcriptional and epigenetic mechanisms of addiction[J]. Nature Reviews Neuroscience,2011,12(11):623-637. doi: 10.1038/nrn3111 [19] 陈绮,梁军成,邓艳萍. 药物成瘾与表观遗传学[J]. 中国药物依赖性杂志,2015,24(1):1-5. [20] Keiichiro M,Mika A,Hiroyuki O. Role of immediate-early genes in synaptic plasticity and neuronal ensembles underlying the memory trace[J]. Frontiers in Molecular Neuroscience,2016,8(1):78. [21] Steward O,Wallace C S,Lyford G L,et al. Synaptic activation causes the mRNA for the IEG Arc to localize selectively near activated postsynaptic sites on dendrites[J]. Neuron,1998,21(4):741. [22] Guzowski J F,Mcnaughton B L,Barnes C A. Environment-specific expression of the immediate-early gene Arc in hippocampal neuronal ensembles[J]. Nature Neuroscience,1999,2(12):1120-1124. [23] Reul,Johannes M. Making memories of stressful events:A journey along epigenetic,gene transcription,and signaling pathways[J]. Frontiers in Psychiatry,2014,5(1):5. [24] Bubenikova-Valesova V,Kacer P,Syslova K,et al. Prenatal methamphetamine exposure affects the mesolimbic dopaminergic system and behavior in adult offspring[J]. International Journal of Developmental Neuroscience,2009,27(6):525-530. doi: 10.1016/j.ijdevneu.2009.06.012 [25] Itzhak Y,Ergui I,Young J I. Long-term parental methamphetamine exposure of mice influences behavior and hippocampal DNA methylation of the offspring[J]. Molecular Psychiatry,2015,20(2):232. doi: 10.1038/mp.2014.7 -

 点击查看大图
点击查看大图

计量
- 文章访问数: 3532
- HTML全文浏览量: 2162
- PDF下载量: 163
- 被引次数: 0
