

Genetics Analysis in a Congenital Cataract Pedigree Associated with a Missense Mutation in GJA8 gene
-
摘要:
目的 通过使用外显子测序定位分析一先天性白内障家系中GJA8基因致病错义突变。 方法 对2020年6月在昆明医科大学第二附属医院就诊的一个先天性白内障家系全体成员进行详细的临床眼科检查及全身查体。采集先证者及6个亲属外周血并提取基因组DNA,应用全外显子测序筛查可疑致病基因,使用生物信息工具对可疑基因突变进行致病性分析,并对家系全部成员进行Sanger测序验证候选致病突变。 结果 外显子测序及生物信息学分析显示GJA8基因存在一个错义突变c.593G > A,p.R198Q,导致其第198位氨基酸残基由谷氨酰胺取代了原有的脯氨酸。氨基酸保守性分析显示该突变影响的氨基酸在物种间高度保守。在家系全部受检者中进行的Sanger测序结果表明该突变与疾病表型共分离,可以认定该突变是该突变为该家系的致病性突变,系谱分析显示该突变所致先天性白内障呈现常染色体显性遗传。 结论 位于GJA8基因的错义突变c.593G > A,p.R198Q是导致该家系出现先天性白内障的遗传病因,遗传方式为常染色体显性遗传。 Abstract:Objective To analyze the clinical manifestations in a congenital cataract pedigree and the genetic etiology was identified using exon sequencing. Methods A family with congenital cataract admitted to the Second Affiliated Hospital of Kunming Medical University in June 2020. A comprehensive ophthalmological examination and physical examination was conducted with the family members. Peripheral blood from proband and 6 relatives were collected, and genomic DNA was extracted. The potential pathogenic genes were screened by whole exome sequencing, pathogenicity analysis of suspected gene mutations was performed using bioinformatics tools, and the mutations were identified using Sanger sequencing in all pedigree members. Results A missense mutation c.593G > A,p.R198Q was identified in GJA8 gene by Exon sequencing and bioinformatics analysis. The GJA8 gene is one of gene family encoding gap junction protein. The amino acids affected by this mutations site are highly conserved between species and the mutation was considered pathogenic according to the Guidelines of the American College of Medical Genetics and Genomics (ACMG). Sanger sequencing results in all in the family showed that the proband and other patients with congenital cataract all carried the mutation, while the mutation was not detected in healthy people in the family, and the mutation was co-separated with the disease phenotype, presenting autosomal dominant inheritance. Conclusions Our study shows that missense mutation c.593G > A,p.R198Q located in the GJA8 gene is the genetic cause of congenital cataract in this family, and the inheritance mode is autosomal dominant inheritance. -
Key words:
- Congenital cataract /
- GJA8 gene /
- Missense mutation /
- Pedigree
-

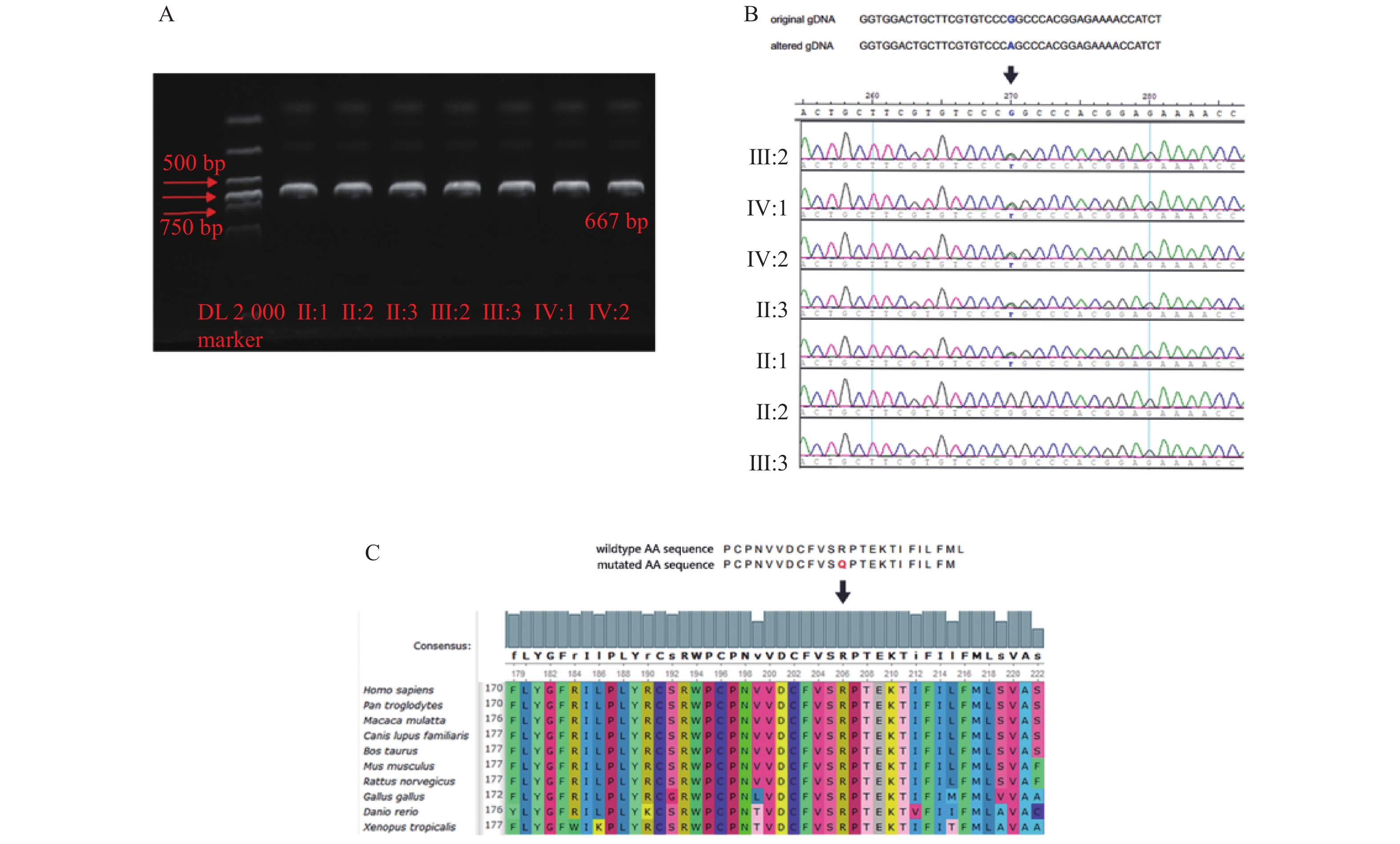
图 1 先天性白内障患者家系图
□:正常男性;○:正常女性;●:女性携带者;■:男性患者;↗:先证者。
Figure 1. Pedigree of congenital cataract patients

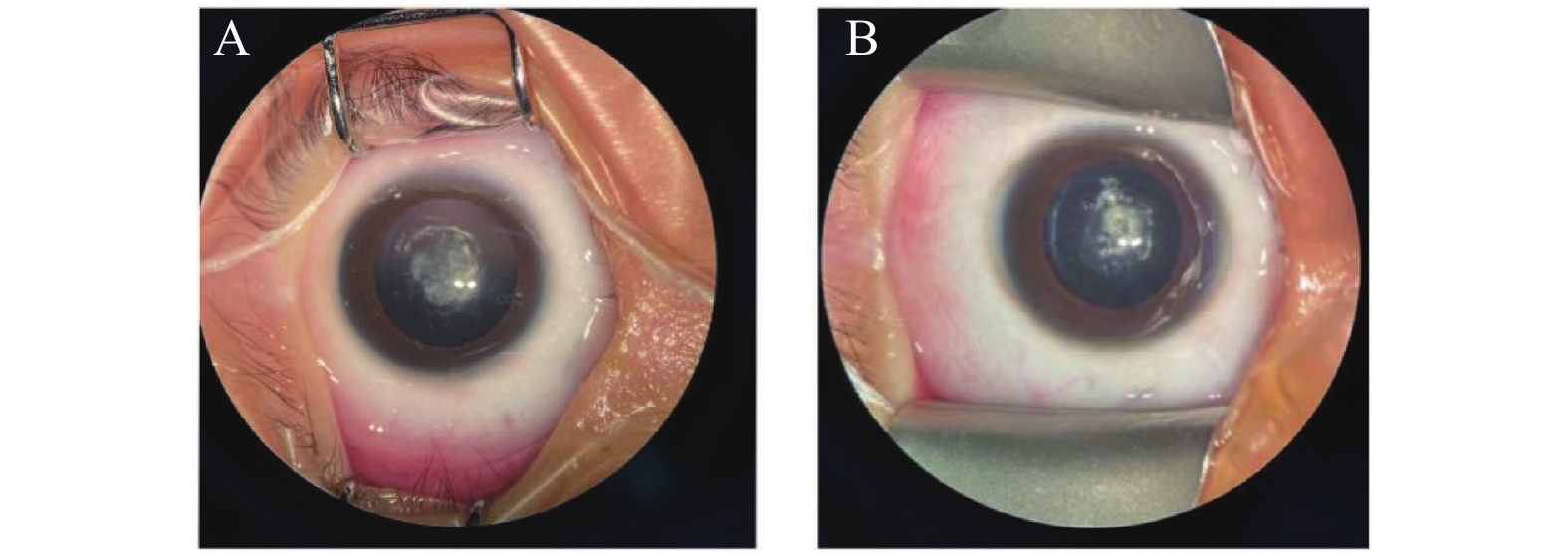
图 2 患者晶状体照片
A:先证者儿子Ⅳ1右眼,晶体后囊膜中央见3×4 mm白色混浊,周边皮质不均匀混浊;B:先证者儿子Ⅳ2右眼,晶体后囊膜中央可见大小约3×4 mm白色混浊,周边皮质不均匀混浊。
Figure 2. Lens photos of the patient
-
[1] Bremond-Gignac D,Daruich A,Robert M P,et al. Recent developments in the management of congenital cataract[J]. Ann Transl Med,2020,8(22):1545. doi: 10.21037/atm-20-3033 [2] Sheeladevi S,Lawrenson J G,Fielder A R,et al. Global prevalence of childhood cataract:A systematic review[J]. Eye (Lond),2016,30(9):1160-1169. doi: 10.1038/eye.2016.156 [3] Lagrèze W A. Treatment of congenital and early childhood cataract[J]. Ophthalmologe,2021,118(Suppl 2):135-144. [4] Li J,Chen X,Yan Y,et al. Molecular genetics of congenital cataracts[J]. Exp Eye Res,2020,191:107872. doi: 10.1016/j.exer.2019.107872 [5] Pichi F,Lembo A,Serafino M,et al. Genetics of congenital cataract[J]. Dev Ophthalmol,2016,57:1-14. [6] Adams D R,Eng C M. Next-generation sequencing to diagnose suspected genetic disorders[J]. N Engl J Med,2018,379(14):1353-1362. doi: 10.1056/NEJMra1711801 [7] Kandaswamy D K,Prakash M V S,Graw J,et al. Application of WES towards molecular investigation of congenital cataracts:Identification of novel alleles and genes in a hospital-based cohort of South India[J]. Int J Mol Sci,2020,21(24):9569. doi: 10.3390/ijms21249569 [8] Li H,Durbin R. Fast and accurate long-read alignment with Burrows-Wheeler transform[J]. Bioinformatics,2010,26(5):589-595. doi: 10.1093/bioinformatics/btp698 [9] Li H,Handsaker B,Wysoker A,et al. The sequence alignment/map format and SAMtools[J]. Bioinformatics,2009,25(16):2078-2079. doi: 10.1093/bioinformatics/btp352 [10] Wang K,Li M,Hakonarson H. ANNOVAR:functional annotation of genetic variants from high-throughput sequencing data[J]. Nucleic Acids Res,2010,38(16):e164. doi: 10.1093/nar/gkq603 [11] Lim M E,Buckley E G,Prakalapakorn S G. Update on congenital cataract surgery management[J]. Curr Opin Ophthalmol,2017,28(1):87-92. doi: 10.1097/ICU.0000000000000324 [12] Chan W H,Biswas S,Ashworth J L,et al. Congenital and infantile cataract:aetiology and management[J]. Eur J Pediatr,2012,171(4):625-630. doi: 10.1007/s00431-012-1700-1 [13] Louison S,Blanc J,Pallot C,et al. Visual outcomes and complications of congenital cataract surgery[J]. J Fr Ophtalmol,2019,42(4):368-374. doi: 10.1016/j.jfo.2018.10.007 [14] Beyer E C,Berthoud V M. Gap junction gene and protein families:Connexins,innexins,and pannexins[J]. Biochim Biophys Acta Biomembr,2018,1860(1):5-8. doi: 10.1016/j.bbamem.2017.05.016 [15] Xin L,Bai D. Functional roles of the amino terminal domain in determining biophysical properties of Cx50 gap junction channels[J]. Front Physiol,2013,4:373. [16] Berthoud V M,Ngezahayo A. Ngezahayo A. Focus on lens connexins[J]. BMC Cell Biol,2017,18(Suppl 1):6. [17] 王秋菊,沈亦平,邬玲仟,等. 遗传变异分类标准与指南[J]. 中国科学:生命科学,2017,47(6):668-688. [18] 王中英,刘善贺,郑贵倩,等. 先天性白内障一患病家系的突变基因筛查[J]. 中国当代医药,2020,27(5):4-6+11+253-254. doi: 10.3969/j.issn.1674-4721.2020.05.003 [19] Ding N,Chen Z,Song X,et al. Novel mutation of GJA8 in autosomal dominant congenital cataracts[J]. Ann Transl Med,2020,8(18):1127. doi: 10.21037/atm-20-4663 [20] 高小博,马燕,马旭,等. 连接蛋白Cx50 S258F突变体对斑马鱼晶状体发育的影响[J]. 中国计划生育学杂志,2012,20(5):313-317. doi: 10.3969/j.issn.1004-8189.2012.05.007 [21] Li L,Fan D B,Zhao Y T,et al. GJA8 missense mutation disrupts hemichannels and induces cell apoptosis in human lens epithelial cells[J]. Sci Rep,2019,9(1):19157. doi: 10.1038/s41598-019-55549-1 [22] Devi R R,Vijayalakshmi P. Novel mutations in GJA8 associated with autosomal dominant congenital cataract and microcornea[J]. Mol Vis,2006,12(21-22):190-195. [23] Yu X,Ping X,Zhang X,et al. The impact of GJA8 SNPs on susceptibility to age-related cataract[J]. Hum Genet,2018,137(11-12):897-904. doi: 10.1007/s00439-018-1945-5 [24] Ceroni F,Aguilera-Garcia D,Chassaing N,et al. New GJA8 variants and phenotypes highlight its critical role in a broad spectrum of eye anomalies[J]. Hum Genet,2019,138(8-9):1027-1042. doi: 10.1007/s00439-018-1875-2 -

 下载:
下载:
 点击查看大图
点击查看大图

图(3)
计量
- 文章访问数: 2962
- HTML全文浏览量: 1926
- PDF下载量: 38
- 被引次数: 0
