

HLA-G Enhances the Killing Effect of NK Cells on Cervical Cancer Cells by Promoting the Expression of Activated Receptor
-
摘要:
目的 探索宫颈癌细胞中HLA-G表达差异对NK细胞表面活化性受体的影响。 方法 实时定量PCR(real-time PCR)及免疫印迹法(Western-Blot WB)检测宫颈癌细胞及正常宫颈细胞中 HLA-G的表达差异。利用慢病毒载体构建 HLA-G稳定下调的宫颈癌细胞株,并与 NK 细胞共培养,CCK8法检测共培养后宫颈癌细胞增殖能力的变化,流式细胞术检测宫颈癌细胞凋亡水平。同时,流式细胞术检测共培养后NK细胞表面活化性受体NKG2D、NKP30的表达水平及颗粒素(Granzyme)、穿孔酶(Perforin)的表达差异。 结果 RT-PCR及WB法检测结果显示:与H8细胞相比,C33a、SiHa细胞中HLA-G的表达显著升高(P < 0.05)。CCK8法检测结果显示:HLA-G下调后,C33a及SiHa细胞与NK细胞共培养,C33a及SiHa细胞的增殖能力显著减弱(P < 0.05),同时,C33a及SiHa细胞的凋亡水平显著升高(P < 0.05)。流式细胞术检测结果显示:C33a及SiHa细胞HLA-G下调后,与其共培养的NK表面NKG2D表达显著升高(P < 0.05),与C33a共培养的NK细胞表面NKP30及Granzyme、Perforin表达显著升高(P < 0.05),与SiHa共培养的NK细胞表面NKP30及Granzyme、Perforin表达存在升高趋势,但差异均无统计学意义(P > 0.05)。 结论 HLA-G在宫颈癌细胞中高表达,HLA-G表达升高可促进活化性受体的表达,增强NK细胞对宫颈癌细胞的杀伤作用。 Abstract:Objective To explore the effect of HLA-G expression on NK cell surface activation receptor in cervical cancer cells. Methods Real-time PCR (RT-PCR) and Western Blot (WB) methods were used to examine the expression of HLA-G in cervical cancer cells and normal cervical cells, respectively. The cervical cancer cell lines transfected with lentivirus to silence the expression of HLA-G were co-cultured with NK cells. The proliferation ability of co-cultured cervical cancer cells was measured by the method of CCK8, and the level of apoptosis in cervical cancer cells was measured by the flow cytometry method, as well as the expression of NKG2D and NKP30, Granzyme and Perforin. Results The results of RT-PCR and WB showed that compared with H8 cells, the expressions of HLA-G in C33a and SiHa cells were significantly increased (P < 0.05). Results of CCK8 showed that the proliferation of C33a and SiHa cells was significantly decreased (P < 0.05) when C33a and SiHa cells were co-cultured with NK cells after transfection. The levels of apoptosis in C33a and SiHa cells were significantly increased (P < 0.05).The results of Flow cytometry showed that the expression of NKG2D on NK surface was significantly increased after C33a and SiHa were transfected (P < 0.05). Of C33a, The expression of NKP30 on the surface of NK cells and Granzyme, Perforin were significantly increased (P < 0.05). However, of C33a, the expression of NKP30 on the surface of NK cells and Granzyme, Perforin were increased, but the difference was not statistically significant (P > 0.05). Conclusion The high expression of HLA-G is in cervical cancer cells, and it may enhance the killing effect of NK cells on cervical cancer cells by promoting the expression of activated receptor. -
Key words:
- Cervical cancer /
- HLA-G /
- NK cell /
- Immune escape
-

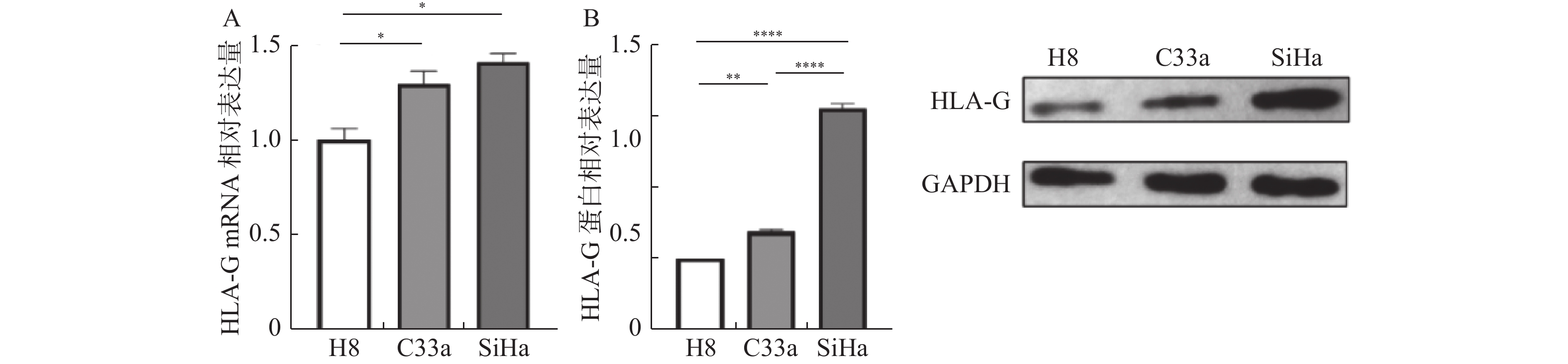
图 2 HLA-G在宫颈癌及正常宫颈上皮细胞系中的表达差异
A:RT-PCR 检测结果;B:WB检测结果。*P < 0.05;**P < 0.01;****P < 0.001。
Figure 2. Difference in HLA-G expression between cervical cancer and normal cervical epithelial cell lines

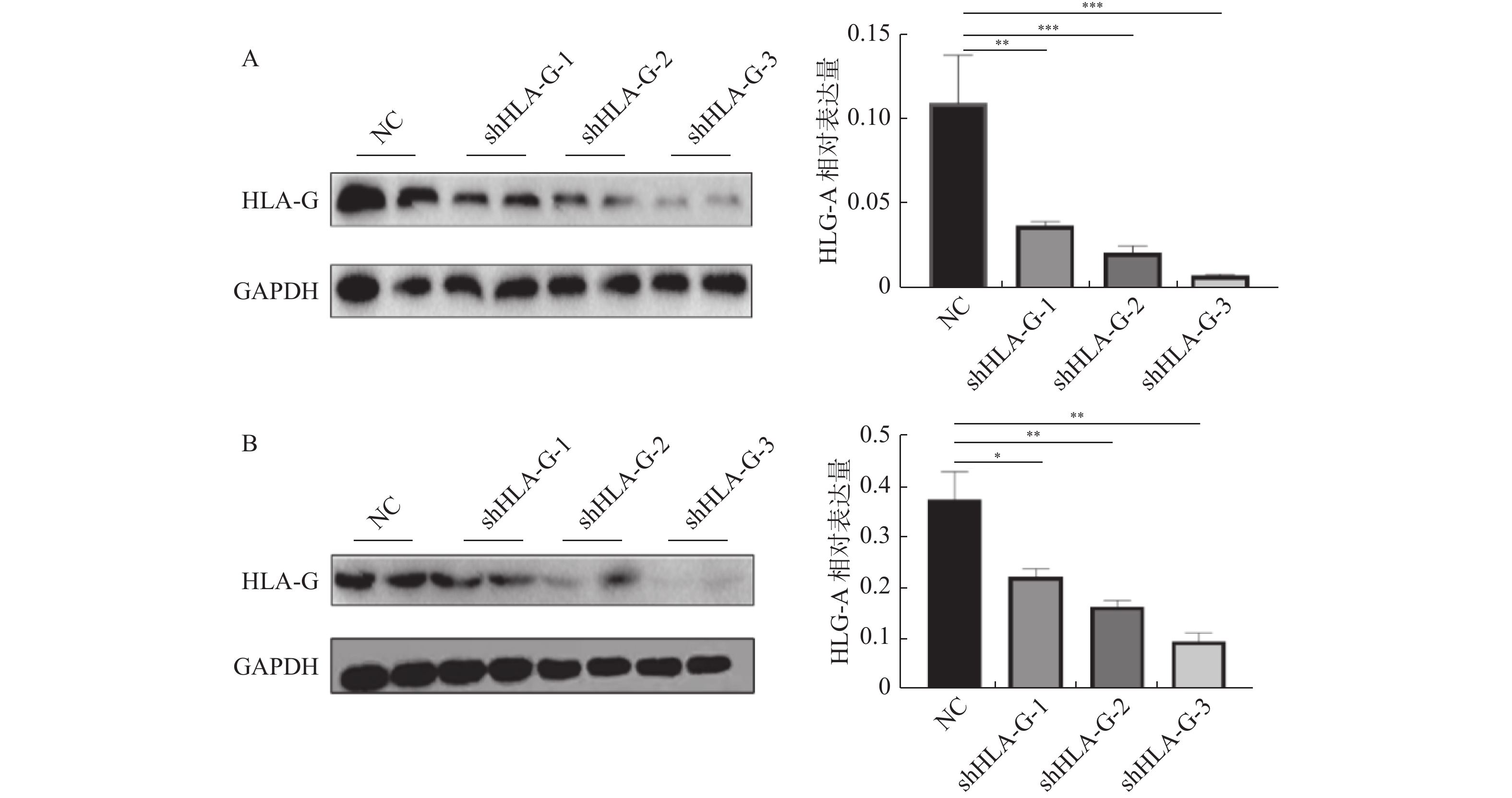
图 3 慢病毒转染宫颈癌细胞系后HLA-G表达情况
A:C33a细胞系;B:SiHa细胞系。*P < 0.05;**P < 0.01;***P < 0.001。
Figure 3. HLA-G expression after transfection in cervical cancer cell lines

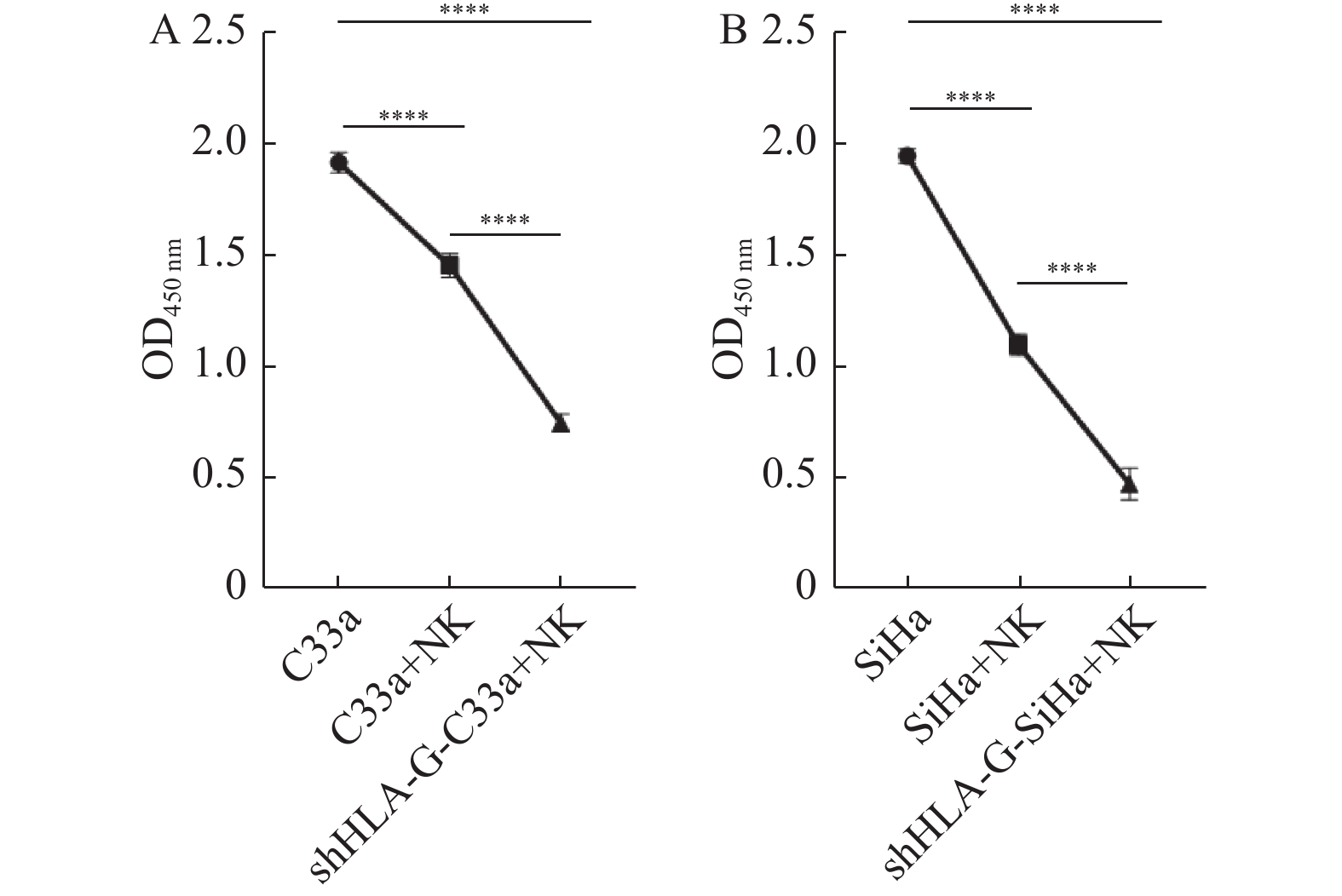
图 4 CCK8法检测各组宫颈癌细胞增殖能力
A:C33a细胞株;B:SiHa细胞株。****P < 0.0001。
Figure 4. CCK8 was used to detect the proliferation of cervical cancer cells

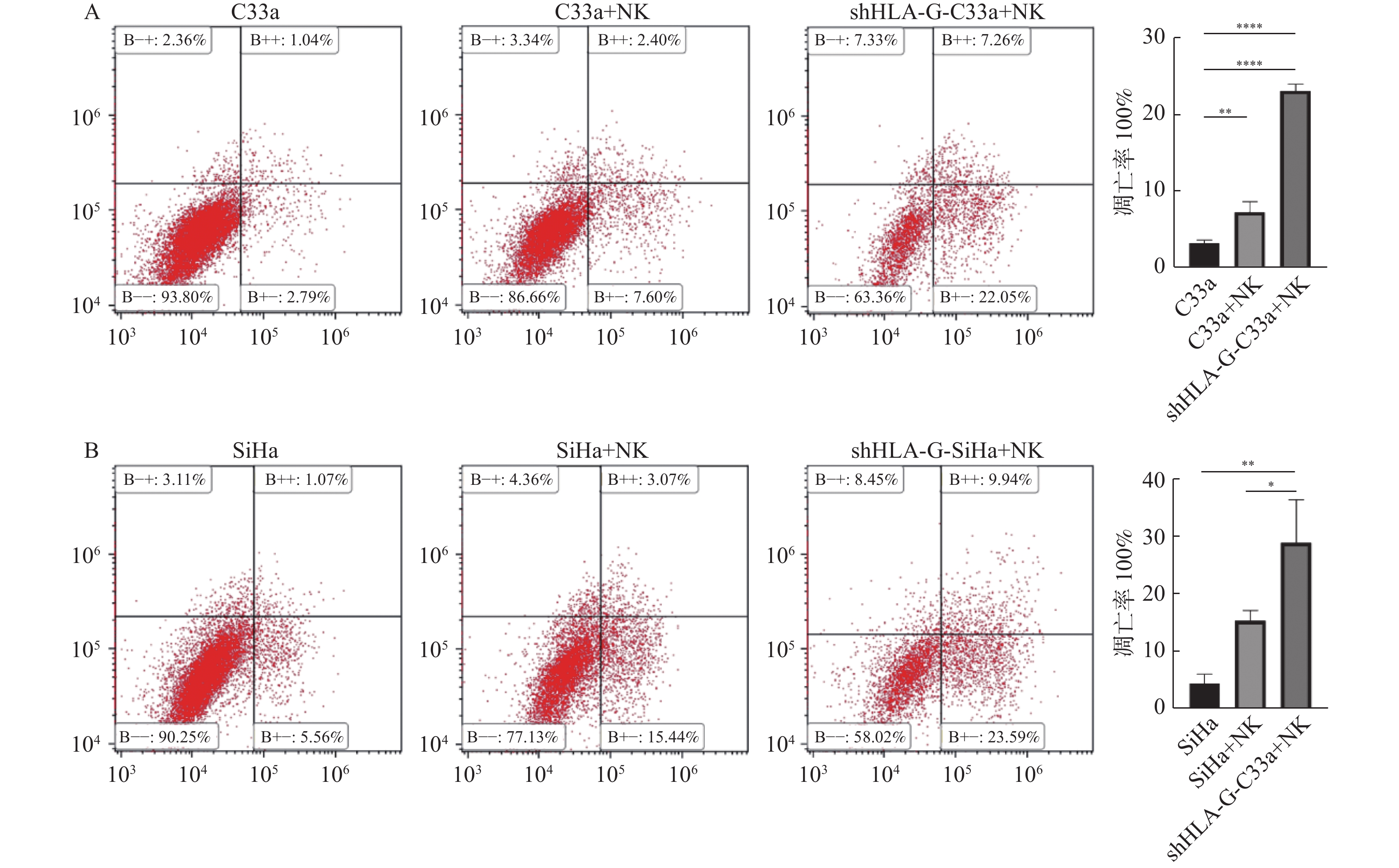
图 5 HLA-G下调后NK细胞与C33a及SiHa细胞共培养,肿瘤细胞凋亡程度
A:C33a 细胞;B:SiHa 细胞。*P < 0.05;**P < 0.01;****P < 0.0001。
Figure 5. After HLA-G transfection,NK cells were co-cultured with C33a and SiHa cells,detection the degree of apoptosis of tumor cells

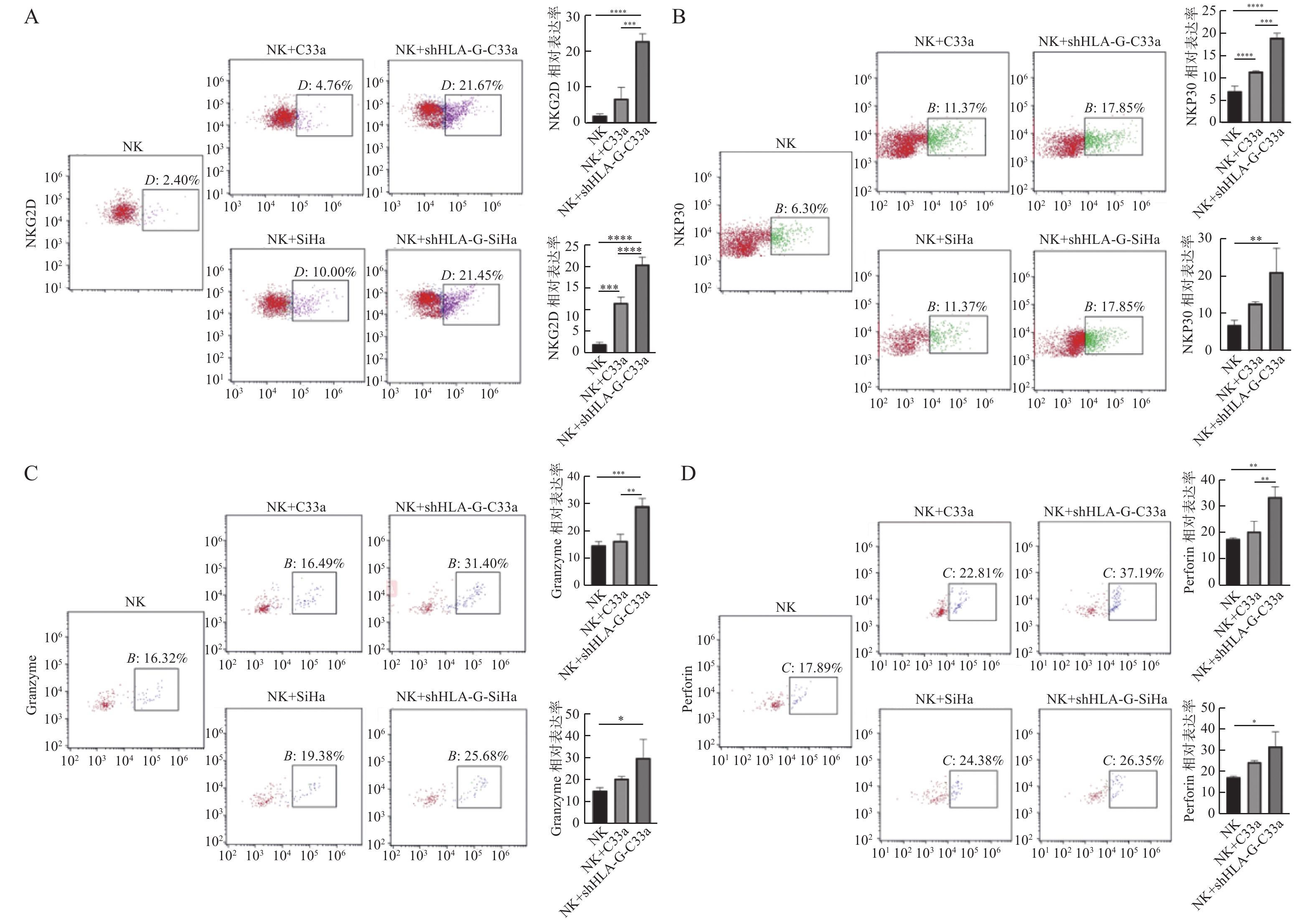
图 6 HLA-G下调后,NK细胞与宫颈癌细胞共培养,NK细胞表面激活性受体及杀伤介质的表达
A:NKG2D;B:NKP30;C:Granzyme;D:Perforin。*P < 0.05;**P < 0.01;****P < 0.0001。
Figure 6. After HLA-G transfection,NK cells were co-cultured with cervical cancer cells,the expression of NK cell surface activating receptor and killing agent was observed
表 1 RT-PCR引物序列
Table 1. RT-PCR primer sequence
基因 引物序列 HLA-G F:GAGAGGAGCAGAGATACATGTG
R:TCAATCTGAGCTCTTCTTCCTCGAPDH F:GAGAAGGCTGGGGCTCATTTGC
R:GCTGATGATCTTGAGGCTGTTGTC 下载: 导出CSV
下载: 导出CSV
-
[1] Buskwofie A,David-West G,Clare C A. A review of cervical cancer:Incidence and disparities[J]. J Natl Med Assoc.,2020,112(2):229-232. [2] 李道娟,师金,靳晶,等. 宫颈癌的流行病学趋势[J]. 中华肿瘤杂志,2021,43(9):912-916. doi: 10.3760/cma.j.cn112152-20190904-00573 [3] Krijgsman D,Roelands J,Hendrickx W,et al. HLA-G:A new immune checkpoint in cancer?[J]. International Journal of Molecular Sciences,2020,21(12):4528. doi: 10.3390/ijms21124528 [4] Jan C I,Huang S W,Canoll P,et al. Targeting human leukocyte antigen G with chimeric antigen receptors of natural killer cells convert immunosuppression to ablate solid tumors[J]. J Immunother Cancer,2021,9(10):e003050. [5] Martín-Villa José Manuel,Vaquero-Yuste Christian,Molina-Alejandre Marta,et al. HLA-G:Too much or too little? role in cancer and autoimmune disease.[J]. Frontiers in Immunology,2022,27(13):796054. [6] 李翠翠,陈梦梦,邹伟,等. 基于NK细胞的免疫疗法在癌症治疗中的应用及其研究进展[J]. 生物学通报,2021,56(12):1-4. doi: 10.3969/j.issn.0006-3193.2021.12.001 [7] Pazina T,Shemesh A,Brusilovsky M,et al. Regulation of the functions of natural cytotoxicity receptors by interactions with diverse ligands and alterations in splice variant expression[J]. Front Immunol,2017,30(8):369. [8] 刘露,陈肖,金慧敏,等. 急性髓系白血病患者NK细胞表达及功能变化研究[J]. 中国实验血液学杂志,2022,30(1):49-55. [9] Ailayangiri S,Altvater B,Spurny C,et al. Targeting Ewing sarcoma with activated and GD2-specific chimeric antigen receptor-engineered human NK cells induces upregulation of immune-inhibitory HLA-G[J]. Oncoimmunology,2016,6(1):e1250050. [10] 李静博,周美英,许惠惠,等. HLA-G与母胎免疫耐受[J]. 医学研究杂志,2017,46(01):12-14. [11] Shen X,Wang P,et al. Correlation between human leukocyte antigen-G expression and clinical parameters in oral squamous cell carcinoma.[J]. Indian Journal of Cancer,2018,55(4):340-343. doi: 10.4103/ijc.IJC_602_17 [12] Zeestraten E,Reimers M S,Saadatmand S,et al. Combined analysis of HLA class I,HLA-E and HLA-G predicts prognosis in colon cancer patients[J]. British Journal of Cancer,2014,110(2):459-468. doi: 10.1038/bjc.2013.696 [13] E M D Kruijf,Sajet A,Nes J,et al. HLA-E and HLA-G expression in classical HLA class I-negative tumors is of prognostic value for clinical outcome of early breast cancer patients[J]. Journal of Immunology,2010,8(12):7452-7459. [14] 陈丽艳,姜继勇,刘莎,于达. 宫颈癌患者高危型HPV感染状况及与血清HLA-G、TGF-β1表达的相关性[J]. 临床和实验医学杂志,2022,21(03):295-299. doi: 10.3969/j.issn.1671-4695.2022.03.020 [15] Dong D D,Yang H,Li K,et al. Human leukocyte antigen-G (HLA-G) expression in cervical lesions:association with cancer progression,HPV 16/18 infection,and host immune response[J]. Reprod Sci,2010,17(8):718-723. doi: 10.1177/1933719110369183 [16] 赵桂增,张晨光. 人类白细胞抗原G参与免疫调节研究进展[J]. 中国免疫学杂志,2020,36(15):1913-1916. doi: 10.3969/j.issn.1000-484X.2020.15.025 [17] Wang L, Dou M, Ma Q, et al. Chimeric antigen receptor (CAR)-modified NK cells against cancer: Opportunities and challenges[J]. Int Immunopharmacol. 2019, 74(期?): 105695.Wang L,Dou M,Ma Q,et al. Chimeric antigen receptor (CAR)-modified NK cells against cancer:Opportunities and challenges[J]. Int Immunopharmacol,2019,74:105695. [18] Guerra N,Tan Y X,Joncker N T,et al. NKG2D-deficient mice are defective in tumor surveillance in models of spontaneous malignancy[J]. Immunity,2008,28(4):571-80. [19] 徐文鑫,张临友. NKG2D与NKG2DL在肺癌免疫调节中的作用及临床意义[J]. 国际免疫学杂志,2012,35(1):39-43. doi: 10.3760/cma.j.issn.1673-4394.2012.01.010 [20] Pende D,Parolini S,Pessino A,et al. Identification and molecular characterization of Nkp30,a novel triggering receptor involved in natural cytotoxicity mediated by human natural killer cells[J]. Journal of Experimental Medicine,1999,190(10):1505-1516. doi: 10.1084/jem.190.10.1505 [21] Pinheiro P F,Justino G C,Marques M M. NKp30 - A prospective target for new cancer immunotherapy strategies[J]. Br J Pharmacol,2020,177(20):4563-4580. [22] Banu N,Riera-Leal A,Haramati J,et al. B7-H6,an immunoligand for the natural killer cell activating receptor NKp30,reveals inhibitory effects on cell proliferation and migration,but not apoptosis,in cervical cancer derived-cell lines[J]. BMC Cancer,2020,20(1):1083. [23] Prager I,Liesche C,van Ooijen H,et al. NK cells switch from granzyme B to death receptor-mediated cytotoxicity during serial killing[J]. J Exp Med,2019,216(9):2113-2127. -

 点击查看大图
点击查看大图
计量
- 文章访问数: 3616
- HTML全文浏览量: 2093
- PDF下载量: 40
- 被引次数: 0
