

Effects of miR-208a on Myocardial Ischemia-reperfusion Injury by Regulating the Expression of QKI5 in Rats
-
摘要:
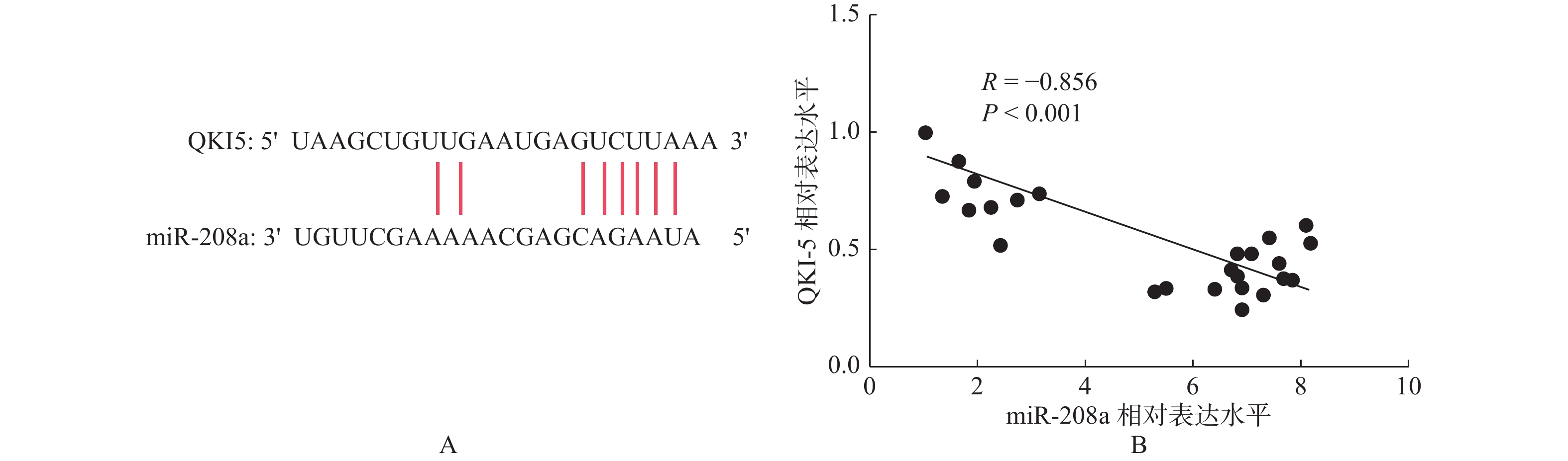
目的 研究miR-208a调控QKI5表达对大鼠心肌缺血再灌注损伤的影响及作用机制。 方法 将雄性SD大鼠32只随机选择分为假手术组(Sham组)、缺血再灌注组(I/R组)、I/R+antagomir阴性对照组(I/R+anta-miR-NC组)和I/R+miR-208a antagomir 组 (I/R+anta-miR-208a组),构建大鼠心肌I/R模型后检测各组大鼠心脏左室射血分数(LVEF)、左室短轴缩短率(LVFS)和左室每搏量(SV);ELISA法检测血清肌酐激酶同工酶(CK-MB)和心肌肌钙蛋白(cTnI)以及IL-1β、IL-6和肿瘤坏死因子(TNF-α)水平;采用HE染色观察各组心肌组织病理变化,采用TUNEL染色检测心肌组织细胞凋亡;RT-PCR检测心肌组织miR-208a和QKI5 mRNA表达,Western blot检测心肌组织QKI5蛋白和凋亡蛋白Caspase-3的相对表达水平。 结果 与Sham组比较,I/R组中LVEF、LVFS、SV值降低,心肌细胞凋亡率升高,血清中CK-MB、cTnI和IL-1β、IL-6、TNF-α水平及心肌组织中miR-208a和Caspase-3表达升高,而QKI5表达水平降低(P < 0.05);与I/R+anta-miR-NC组比较,I/R+anta-miR-208a组中LVEF、LVFS、SV值升高,心肌细胞凋亡率降低,血清中CK-MB、cTnI和IL-1β、IL-6、TNF-α及心肌组织中miR-208a和Caspase-3表达水平降低,而QKI5表达升高(P < 0.05)。相关性分析发现miR-208a与QKI5表达呈负相关。 结论 抑制miR-208a可能通过调控QKI5表达减轻心肌缺血再灌注损伤中炎症反应,发挥心肌保护作用。 -
关键词:
- 心肌缺血再灌注损伤 /
- 微小RNA-208a /
- QKI5蛋白 /
- 相关性
Abstract:Objective To investigate the effect and mechanism of miR-208a on myocardial injury induced by myocardial ischemia reperfusion by regulating the expression of QKI5 in rats. Methods Thirty-two male SD rats were randomly divided into sham group, I/R group, I/R+anta-miR-NC group and I/R+anta-miR-208a group. The rat model of myocardial I/R was established, and the LVEF, LVFS and SV were detected in each group. The levels of CK-MB, cTnI, IL-1β, IL-6, TNF-αwere measured by ELISA. HE staining was used to observe the pathological changes of myocardial tissue in each group, and TUNEL staining was used to detect myocardial cell apoptosis.RT-PCR was used to detect the expression of miR-208a and QKI5 mRNA in myocardial tissue, and Western blot was used to detect the realative expression levels of QKI5 protein and Caspase-3 in myocardial tissue. Results Compared with the Sham group, the levels of LVEF, LVFS, SV in the I/R group were decreased, while the apoptosis rate of cardiomyocytes was increased, and the levels of CK-MB, cTnI, IL-1β, IL-6, TNF-α in serum and the expressions of miR-208a and Caspase-3 in the myocardial tissue were increased, and the expressions of QKI5 were decreased (P < 0.05). Compared with the I/R+anta-miR-NC group, the levels of LVEF, LVFS, SV in the I/R+anta-miR-208a group were increased, while the apoptosis rate of cardiomyocytes was decreased, and the levels of CK-MB, cTnI, IL-1β, IL-6, TNF-α in serum and the expressions of miR-208a and Caspase-3 in the myocardial tissue were decreased, and the expressions of QKI5 were increased (P < 0.05). Correlation analysis found that the expression of miR-208a and QKI5 had negative correlation in the I/R rats. Conclusion Inhibition of miR-208a may alleviate the inflammatory response to exert myocardial protective function in myocardial ischemia-reperfusion injury by regulating the expression of QKI5. -
Key words:
- Myocardial ischemia-reperfusion injury /
- miR-208a /
- QKI5 protein /
- Correlation
-
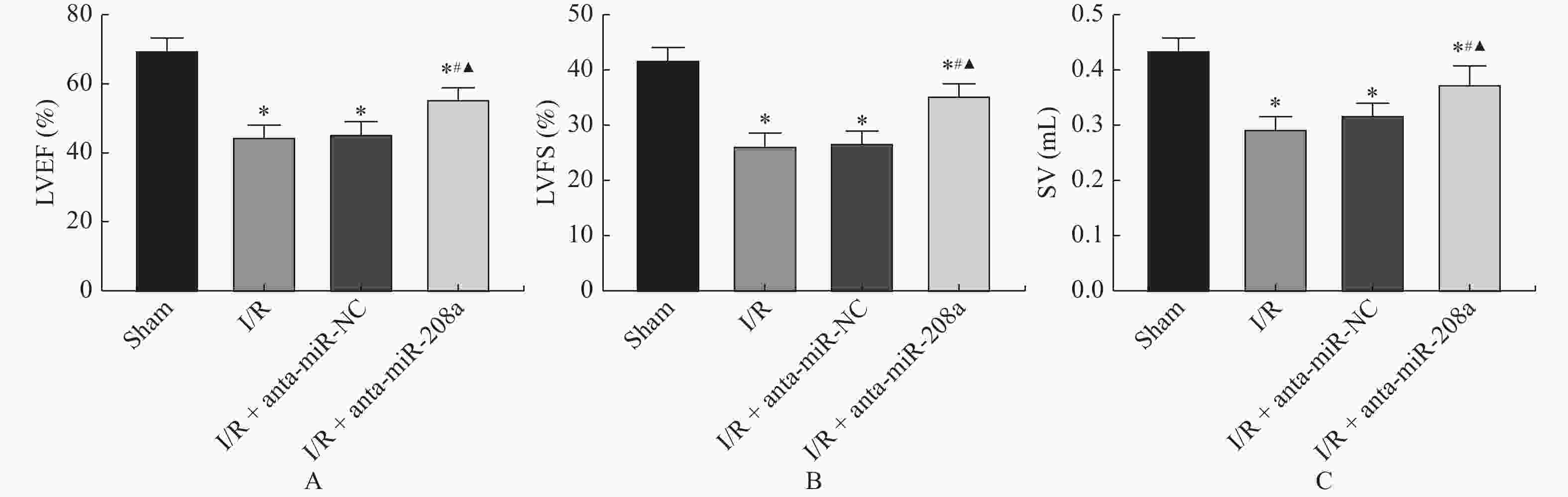
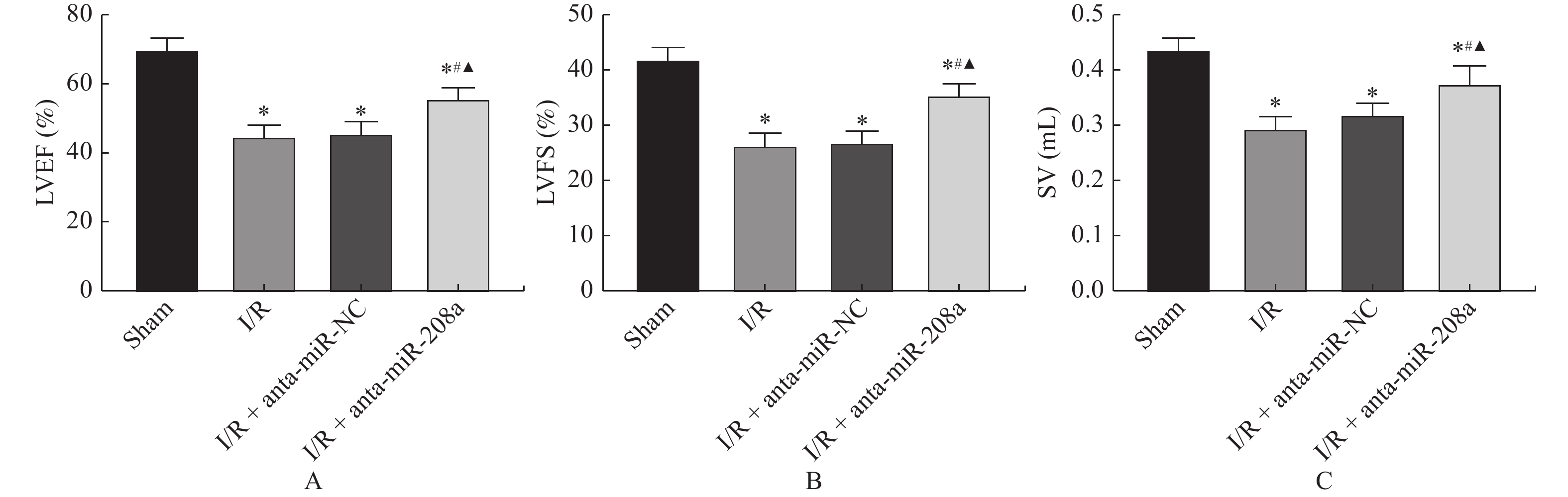
图 1 miR-208a对大鼠心脏功能的影响
A:LVEF值;B:FS值;C:SV值。与Sham组相比,*P < 0.05;与I/R组相比,#P < 0.05;与anta-miR-NC组相比,▲P < 0.05。
Figure 1. Effect of anta-miR-208a on cardiac function in rats(
$\bar x \pm s $ ,n = 8)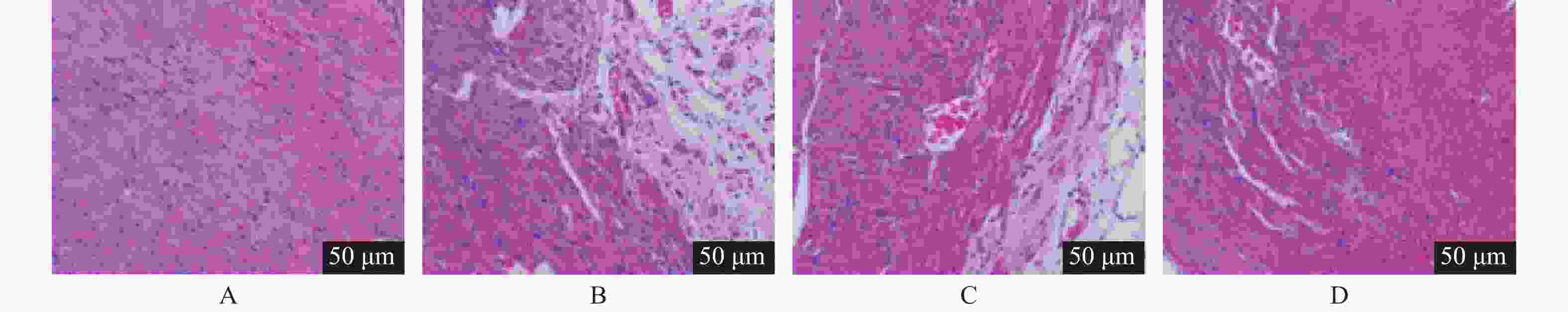
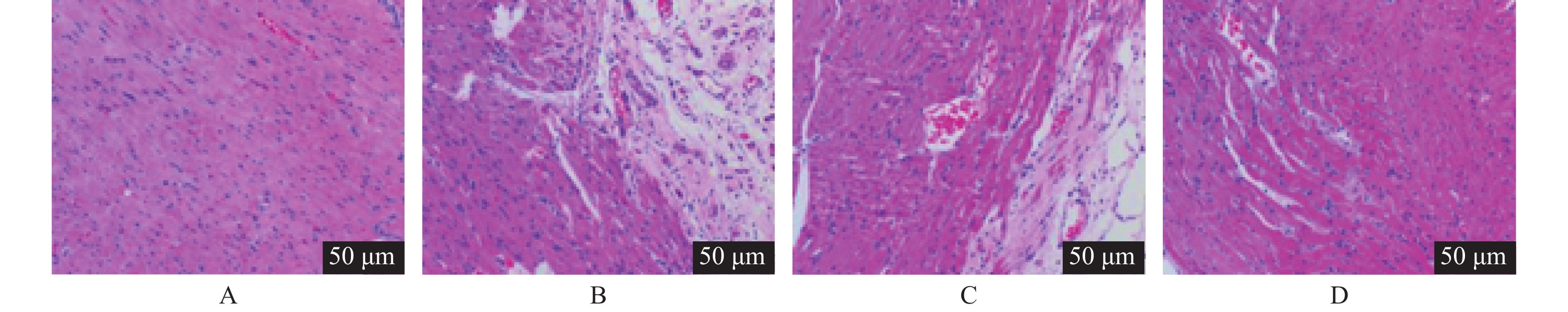
图 2 各组心肌组织病理变化(HE ×200)
A:Sham组;B:I/R组;C:I/R+anta-miR-NC组;D:I/R+anta-miR-208a组。
Figure 2. Pathological changes of myocardial tissue in each group (HE ×200)

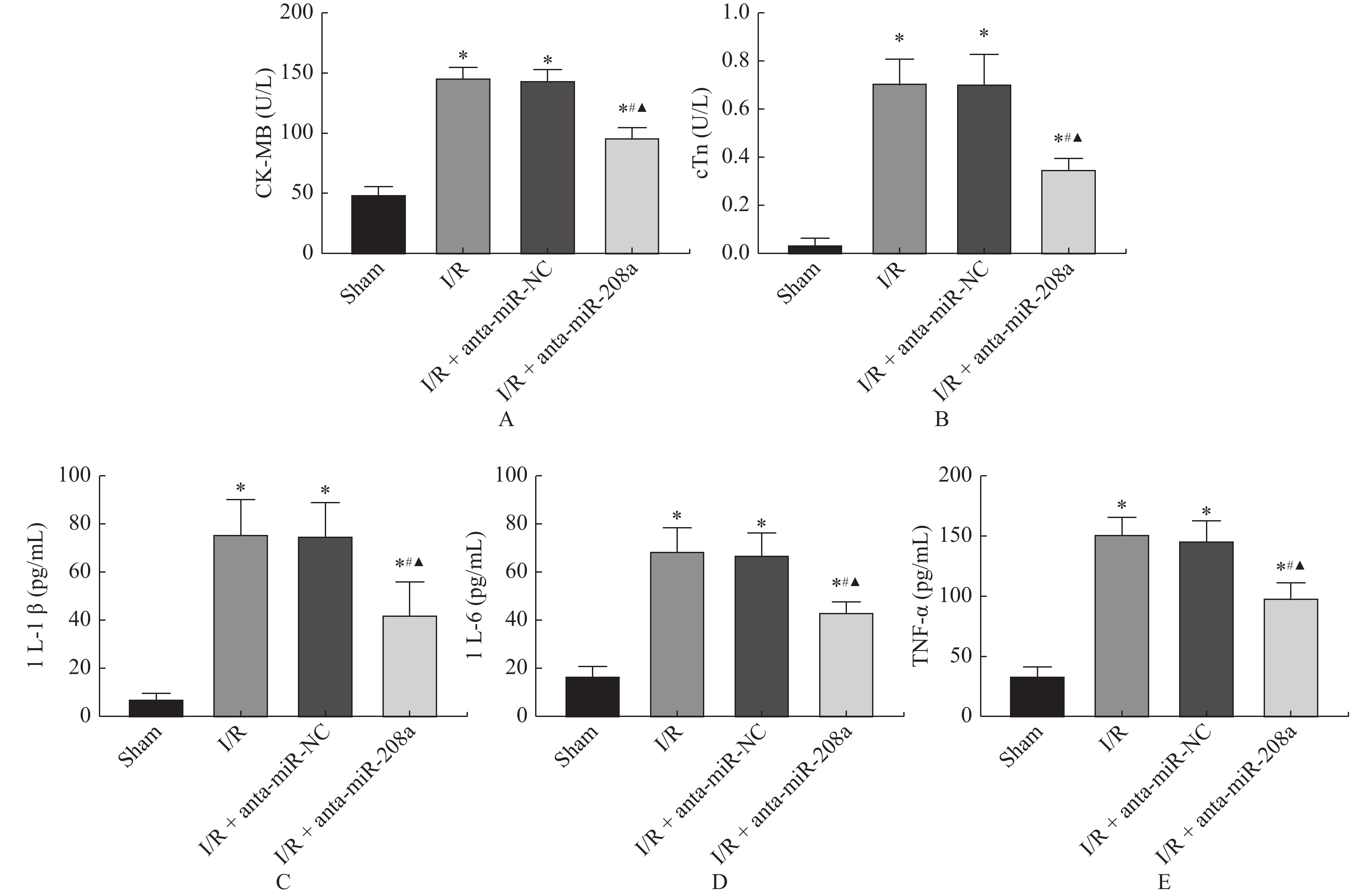
图 3 miR-208a对I/R大鼠心肌损伤标注物和炎性因子的影响
A、B:大鼠血清心肌损伤标志物CK-MB和cTnI水平;C、D、E:大鼠血清炎性因子IL-1β、IL-6和TNF-α水平。与Sham组相比,*P < 0.05;与I/R组相比,#P < 0.05;与anta-miR-NC组相比,▲P < 0.05。
Figure 3. Effects of miR-208a expression on myocardial injury makers and inflammatory factors in I/R rats(
$\bar x \pm s $ ,n = 8)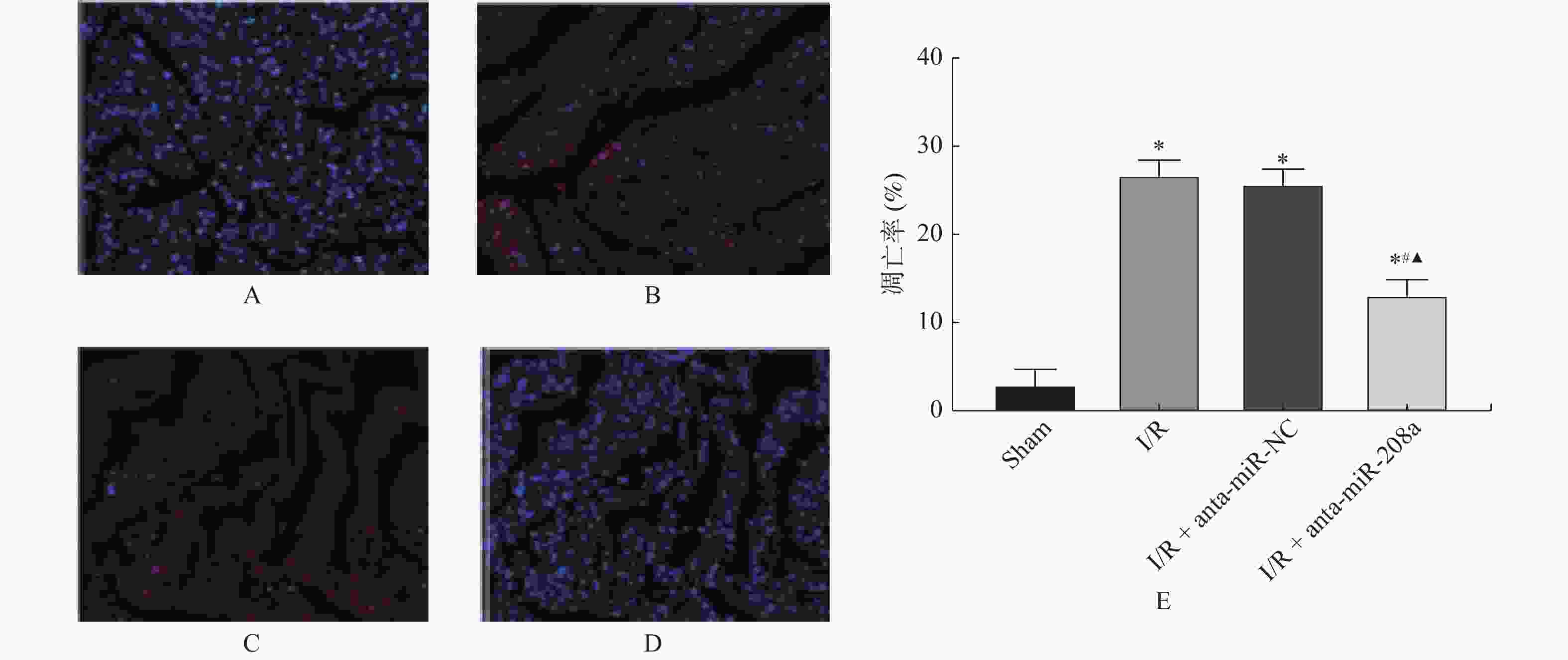
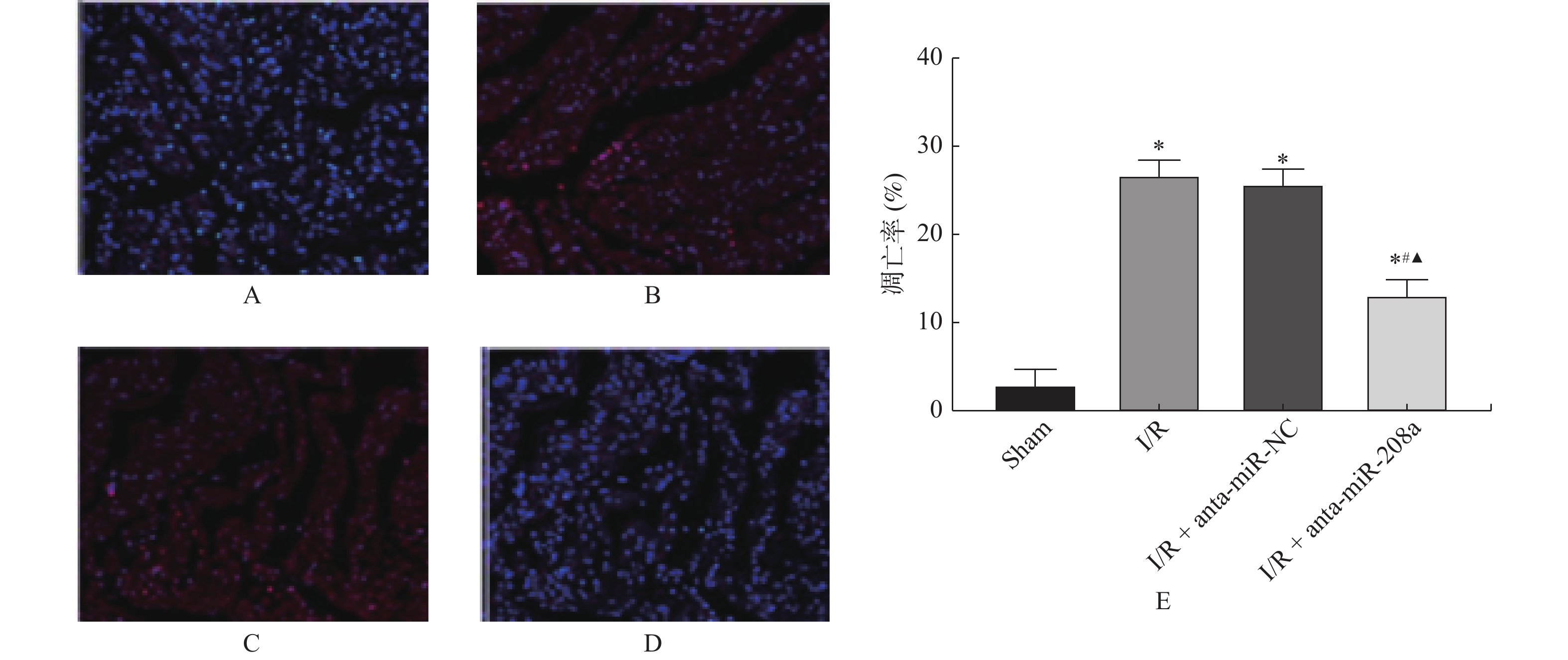
图 4 各组心肌组织细胞凋亡情况(TUNEL×200)
A:Sham组;B:I/R组;C:I/R+anta-miR-NC组;D:I/R+anta-miR-208a组;E:大鼠心肌细胞凋亡率。与Sham组相比,*P < 0.05;与I/R组相比,#P < 0.05;与anta-miR-NC组相比,▲P < 0.05。
Figure 4. Apoptosis of myocardial tissue cells in each group (TUNEL×200)(
$\bar x \pm s $ ,n = 8)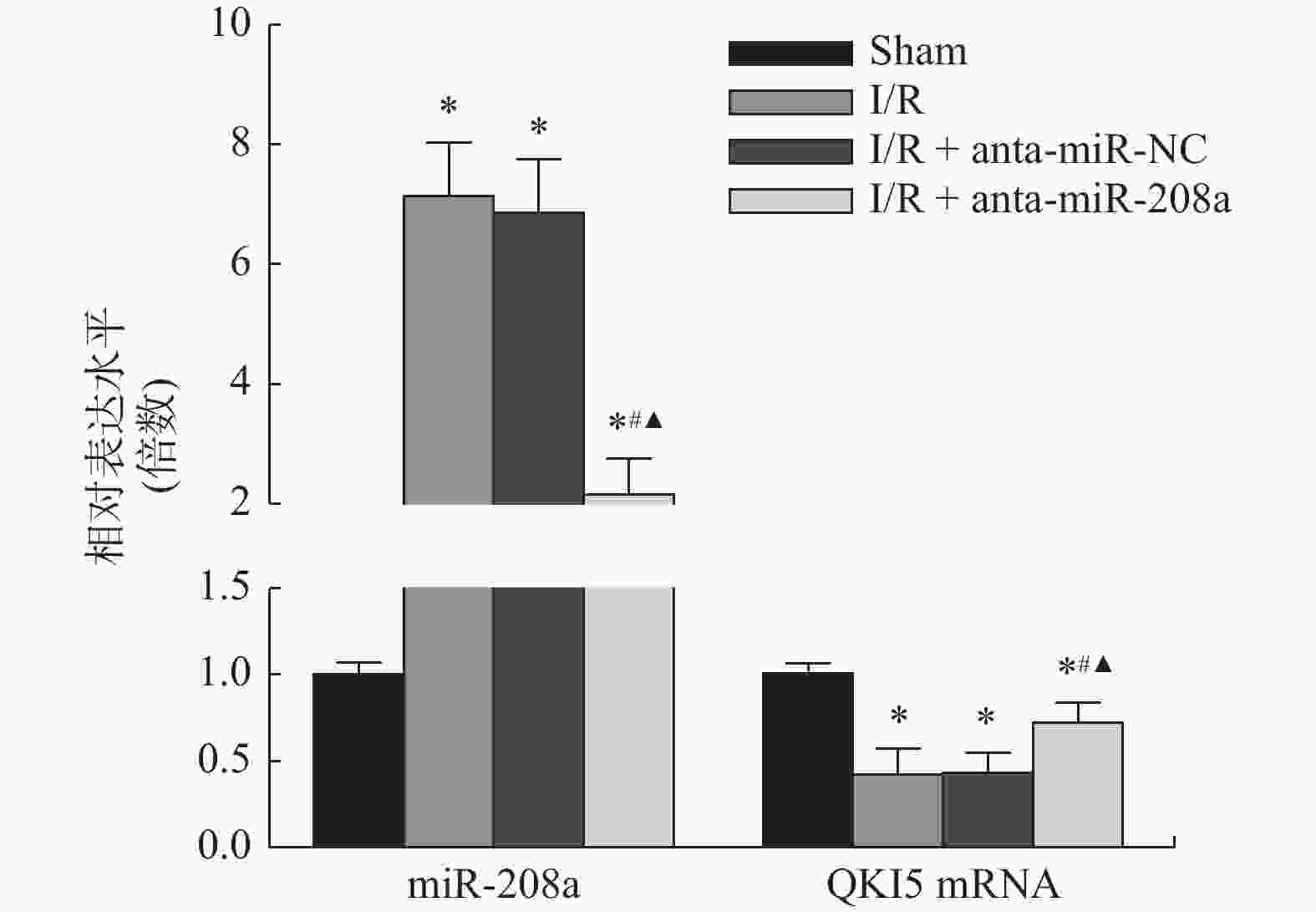
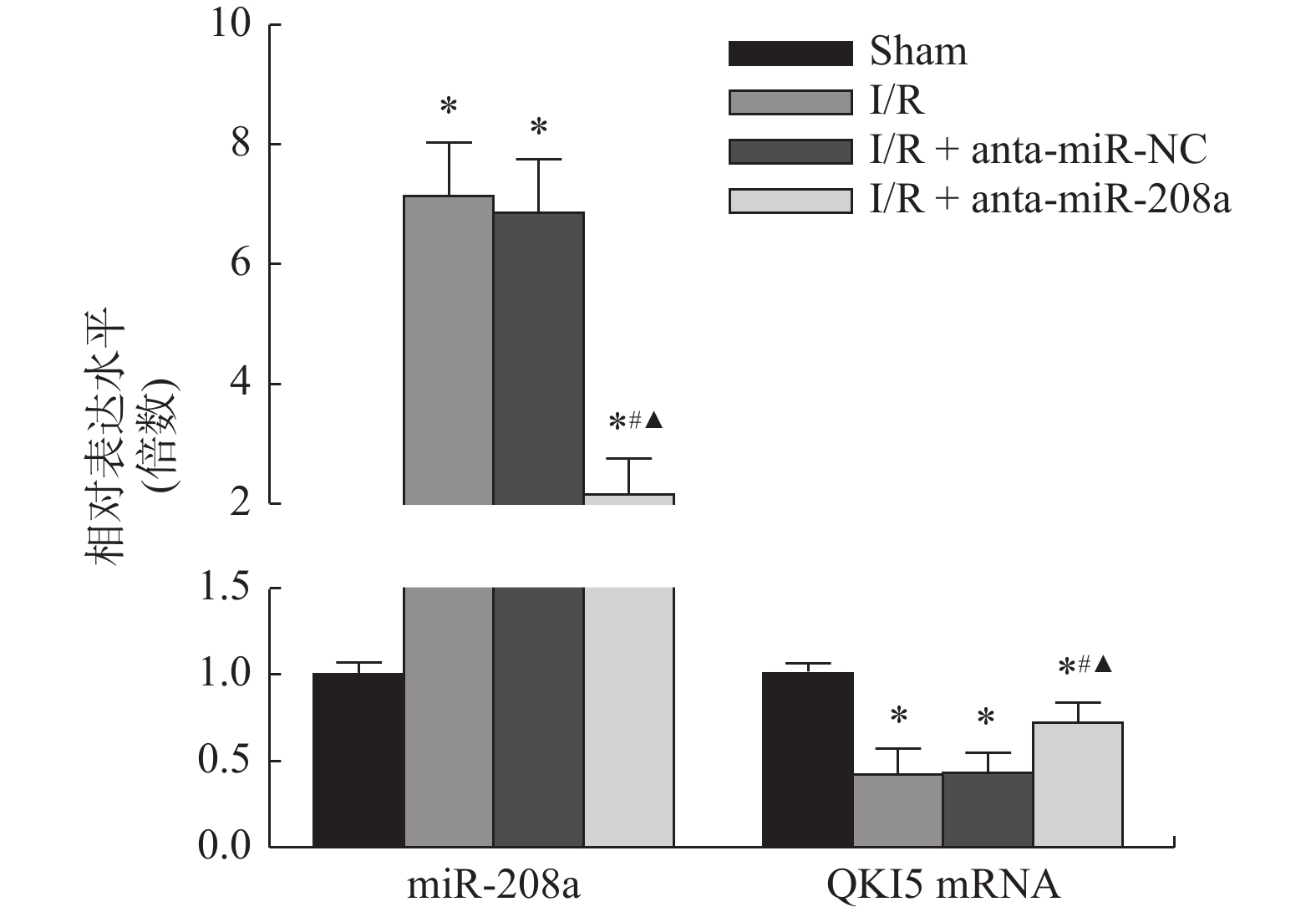
图 5 各组大鼠心肌组织miR-208a和QKI5 mRNA相对表达水平
与Sham组相比,*P < 0.05;与I/R组相比,#P < 0.05;与anta-miR-NC组相比,▲P < 0.05。
Figure 5. Relative expression levels of miR-208a and QKI5 mRNAin myocardial tissue of rats in each group (
$\bar x \pm s $ ,n = 8)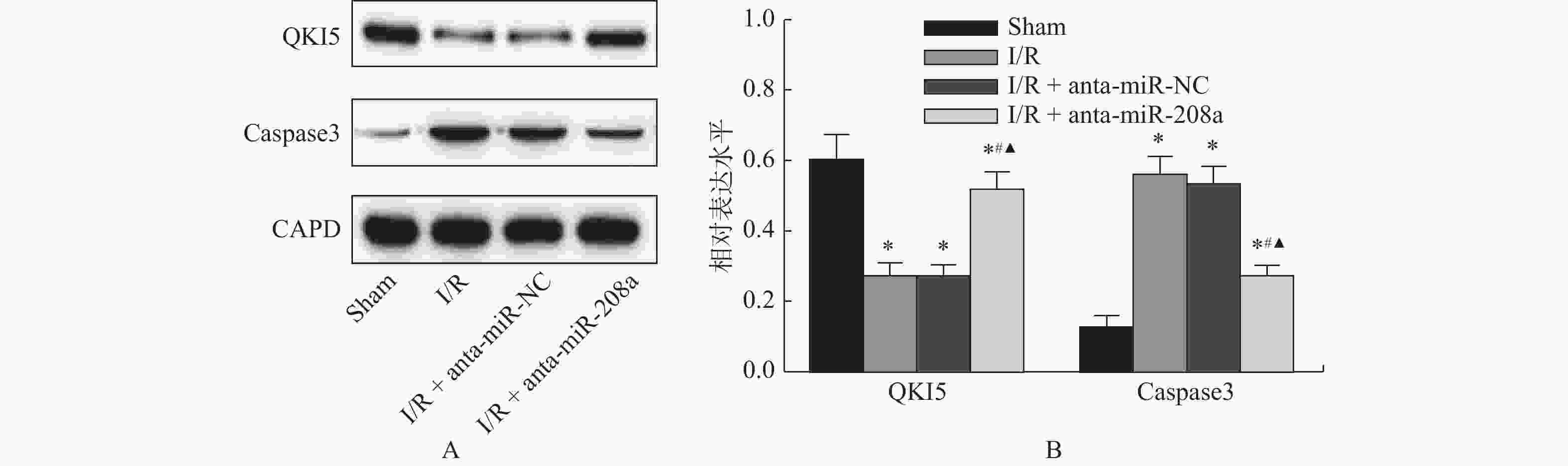
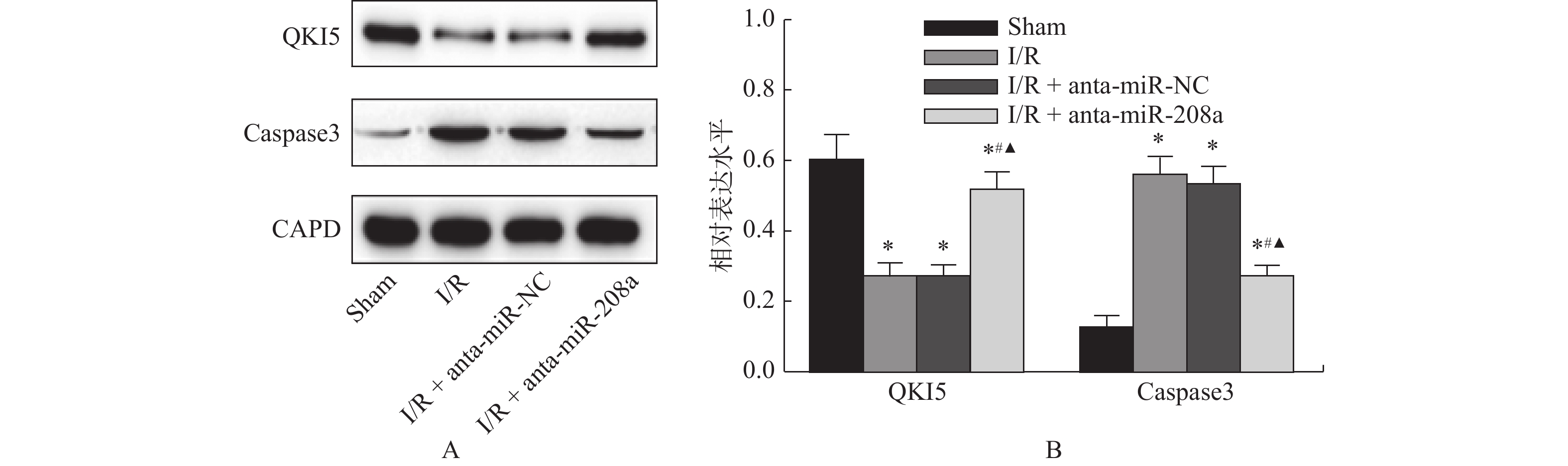
图 6 各组大鼠心肌组织中蛋白的相对表达
A:心肌组织中QKI5和Caspase3的WB图;B:心肌组织中QKI5和Caspase3相对表达量。与Sham组相比,*P < 0.05;与I/R组相比,#P < 0.05;与anta-miR-NC组相比,▲P < 0.05。
Figure 6. Relative expression levels of protein in myocardial tissue of rats in each group(
$\bar x \pm s $ ,n = 8) -
[1] Mokhtari-Zaer A,Marefati N,Atkin S L,et al. The protective role of curcumin in myocardial ischemia-reperfusion injury[J]. Journal of Cellular Physiology,2018,234(1):214-222. [2] Russo I,Penna C,Musso T,et al. Platelets,diabetes and myocardial ischemia/reperfusion injury[J]. Cardiovascular Diabetology,2017,16(1):71. doi: 10.1186/s12933-017-0550-6 [3] 常国楫,顾金松,孙林. MicroRNA在心肌再灌注损伤应用的研究进展[J]. 昆明医科大学学报,2017,38(5):138-143. doi: 10.3969/j.issn.1003-4706.2017.05.031 [4] Tony H,Yu K,Qiutang Z. MicroRNA-208a silencing attenuates doxorubicin induced myocyte apoptosis and cardiac dysfunction[J]. Oxidative Medicine and Cellular Longevity,2015,6(2015):597032. [5] Guo W,Shi X,Liu A,et al. RNA binding protein QKI inhibits the ischemia/reperfusion-induced apoptosis in neonatal cardiomyocytes[J]. Cell Physiol Biochem,2011,28(4):593-602. doi: 10.1159/000335755 [6] Kunecki M,Płazak W,Podolec P,et al. Effects of endogenous cardioprotective mechanisms on ischemia-reperfusion injury[J]. Postepy Hig Med Dosw (Online),2017,71(10):20-31. doi: 10.5604/01.3001.0010.3786 [7] 陈耽,李骊华. miR-208a在大鼠心肌缺血再灌注损伤及缺血后处理中的表达及其作用[J]. 重庆医科大学学报,2015,4(40):501-505. doi: 10.13406/j.cnki.cyxb.000494 [8] Kura B,Szeiffova Bacova B,Kalocayova B,et al. Oxidative Stress-Responsive MicroRNAs in Heart Injury[J]. Int J Mol Sci.,2020,21(1):358. doi: 10.3390/ijms21010358 [9] Wang F,Yuan Y,Yang P,et al. Extracellular vesicles-mediated transfer of miR-208a/b exaggerate hypoxia/reoxygenation injury in cardiomyocytes by reducing QKI expression[J]. Mol Cell Biochem,2017,431(1-2):187-195. doi: 10.1007/s11010-017-2990-4 [10] Yan F,Liu R,Zhuang X,et al. Salidroside attenuates doxorubicin-induced cardiac dysfunction partially through activation of QKI/FoxO1 pathway[J]. J Cardiovasc Transl Res,2021,14(2):355-364. doi: 10.1007/s12265-020-10056-x [11] Guo W,Jiang T,Lian C,et al. QKI deficiency promotes FoxO1 mediated nitrosative stress and endoplasmic reticulum stress contributing to increased vulnerability to ischemic injury in diabetic heart[J]. J Mol Cell Cardiol,2014,75(10):131-140. [12] Al-Salam S,Hashmi S. Myocardial ischemia reperfusion injury:Apoptotic,inflammatory and oxidative stress role of galectin-3[J]. Cell Physiol Biochem,2018,50(3):1123-1139. doi: 10.1159/000494539 [13] Taki J,Wakabayashi H,Inaki A,et al. 14C-Methionine uptake as a potential marker of inflammatory processes after myocardial ischemia and reperfusion[J]. J Nucl Med,2013,54(3):431-436. doi: 10.2967/jnumed.112.112060 [14] Li Y,Li Z,Liu J,et al. miR-190-5p alleviates myocardial ischemia-reperfusion injury by targeting PHLPP1[J]. Disease Markers,2021,2021:8709298. -


 下载:
下载:




 点击查看大图
点击查看大图
计量
- 文章访问数: 4678
- HTML全文浏览量: 2073
- PDF下载量: 36
- 被引次数: 0
