

Effects of Perindopril on Intestinal Microecology in Obese Rats
-
摘要:
目的 研究培哚普利对肥胖大鼠体重、血脂及肠道菌群的影响。 方法 SPF级Wistar大鼠随机分为正常组,模型组,培哚普利低、高剂量组 [0.4、2 mg /(kg·d)] ,连续给药3周后测定各组大鼠血清TC、TG、LDL-C、HDL-C、UA、GLU及粪便中肠道菌群的变化。 结果 与正常组比较,模型组大鼠终体重、体重增重、Lee's 2s 指数升高(P < 0. 01),说明模型建立成功;与模型组比较,培哚普利低、高剂量组大鼠体重明显降低( P < 0.05)。16S rRNA测序结果显示,培哚普利可提高大鼠肠道菌群多样性水平,培哚普利低剂量组OTU总数增加了4.56%;在门和属水平上,培哚普利低剂量组厚壁菌门物种组成显著降低( P < 0. 01) ,拟普雷沃菌属、 NK4A214_group、no_rank_f__Erysipelotrichaceae、norank_f__Eubacterium_coprostanoligenes_group物种组成显著升高(P < 0.05),培哚普利高剂量组放线菌门、 norank_f__Erysipelotrichaceae、Enterorhabdus和葡萄球菌属物种组成显著升高(P < 0. 01)。 结论 培哚普利可降低肥胖大鼠体重,其作用可能与调节大鼠肠道菌群组成有关。 Abstract:Objective To study the effect of perindopril on body weight, blood lipids and intestinal flora of obese rats. Methods SPF Wistar rats were randomly divided into normal group, model group, perindopril low-dose and high-dose groups (0.4, 2 mg/(kg·d)). Changes in LDL-C, HDL-C, UA, GLU and intestinal flora in feces were compared. Results Compared with the normal group, the final body weight, body weight gain and Lee's index of the rats in the model group increased (P < 0.01), indicating that the model was successfully established; compared with the model group, the body weight of the rats in the perindopril low and high dose groups significantly decreased ( P < 0.05, 0.01). The results of 16S rRNA sequencing showed that perindopril could improve the level of intestinal flora diversity in rats, and the total number of OTUs in the perindopril low-dose group increased by 4.56%; at the phylum and genus levels, the perindopril low-dose group The species composition of Firmicutes was significantly decreased (P < 0.01), and the species composition of Alloprevotella, NK4A214_group, no_rank_f__Erysipelotrichaceae, norank_f__Eubacterium_coprostanoligenes_group were significantly increased (P < 0.05, 0.01), and The species compositions of Actinobacteriota, norank_f__Erysipelotrichaceae, Enterorhabdus and Staphylococcus were significantly increased in the perindopril high-dose group (P < 0.01). Conclusion Perindopril can reduce the body weight of obese rats, and its effect may be related to the regulation of intestinal flora composition of rats. -
Key words:
- Perindopril /
- Obesity /
- Rats /
- Intestinal microecology
-

图 1 培哚普利对大鼠肠道菌群的多样性分析
A:韦恩图;B:OTU水平的PLS-DA分析。
Figure 1. Diversity analysis of perindopril on the intestinal flora of rats

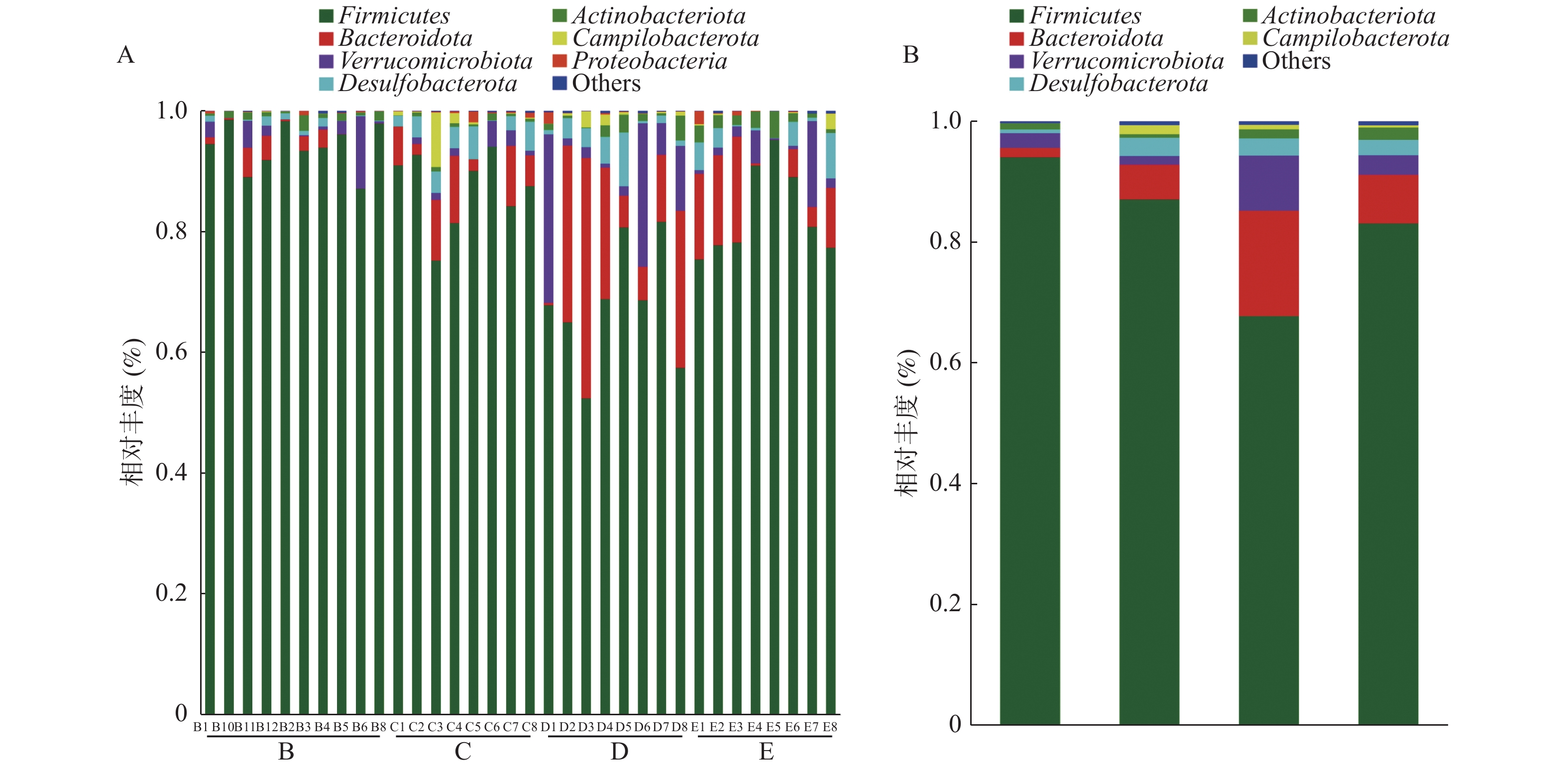
图 2 各组大鼠肠道菌群门水平的优势物种及其相对丰度
A:所有样本;B:分组样本。
Figure 2. Predominant species and their relative abundance at the phylum level of rat gut flora in each group

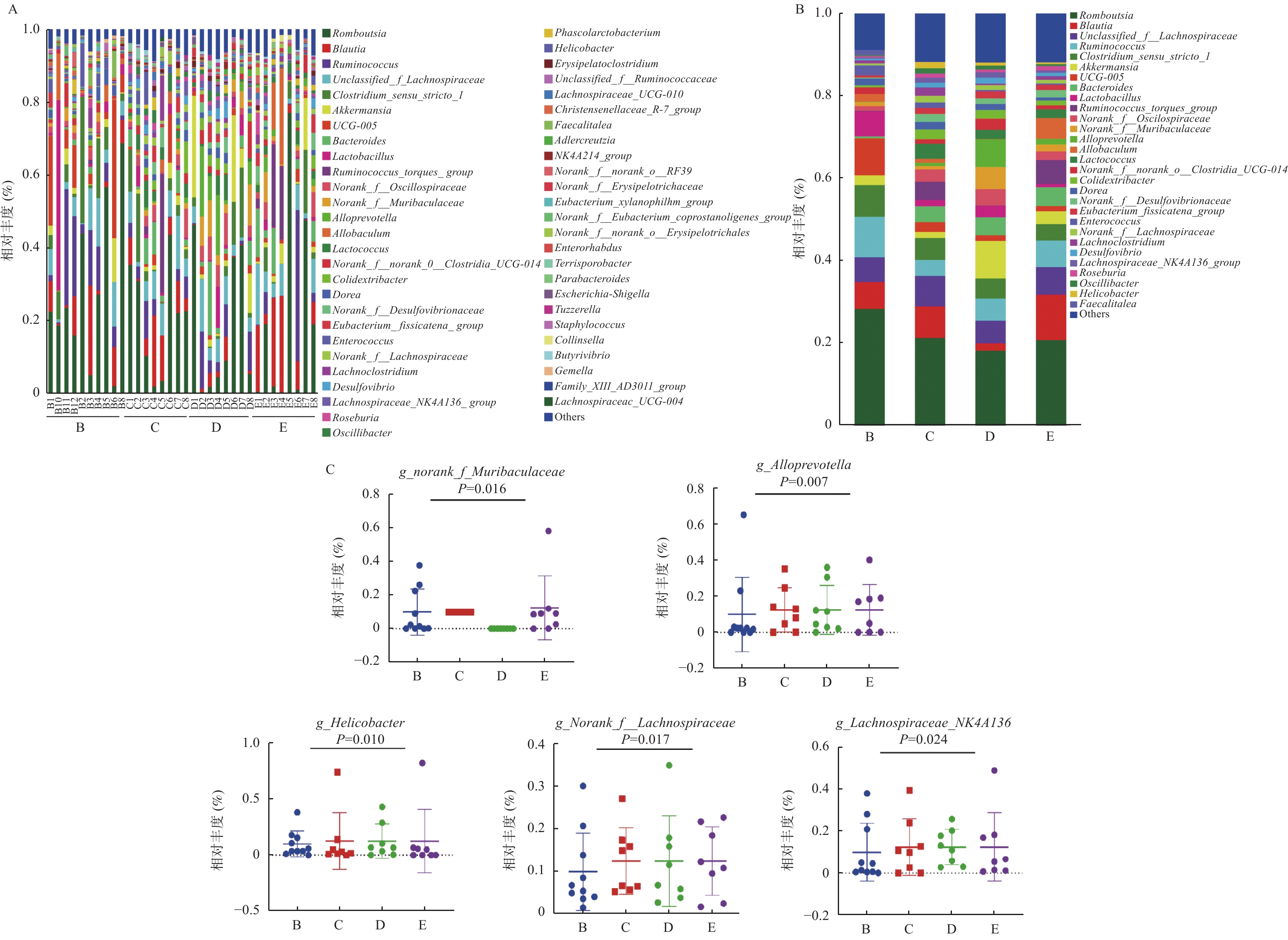
图 3 各组大鼠肠道菌群属水平组成
A:所有样本;B:分组样本;C:各组大鼠粪便属水平比较。
Figure 3. Level composition of the intestinal flora of rats in each group

图 4 大鼠肠道菌群与Lee's 指数、血生化指标Spearman相关性分析
Figure 4. Spearman correlation analysis of rat intestinal flora with Lee's index and blood biochemical index
*P < 0.05,**P < 0.01, ***P < 0.001 。
表 1 培哚普利对高脂饮食大鼠体重和Lee's 指数的影响( $\bar x \pm s $)
Table 1. Effects of perindopril on body weight and Lee's index in high-fat diet rats ( $\bar x \pm s $)
组别 n 初始体重(g) 终体重(g) 体重增重(g) Lee's 指数(g/cm) 正常组 10 127.55 ± 3.45 444.45 ± 22.56 316.90 ± 21.67 2.86 ± 0.05 模型组 8 129.50 ± 6.13 543.90 ± 19.64## 414.40 ± 16.48## 3.06 ± 0.05## 培哚普利低剂量组 8 131.06 ± 4.54 515.50 ± 18.26##* 384.44 ± 18.58##* 2.99 ± 0.06## 培哚普利高剂量组 8 132.85 ± 4.14 480.83 ± 14.05##**@ 347.98 ± 13.26#**@@ 3.00 ± 0.08## 与正常组比较,#P < 0.05, ##P < 0.01;与模型组比较, *P < 0.05, **P < 0.01;与培哚普利低剂量组比较, @P < 0.05, @@P < 0.01。  下载: 导出CSV
下载: 导出CSV
表 2 培哚普利对对高脂饮食大鼠血生化指标的影响[( $\bar x \pm s $),mmol /L]
Table 2. Effects of perindopril on blood biochemical indexes in high-fat diet rats [( $\bar x \pm s $),mmol /L)
组别 n TC TG LDL-C HDL-C UA GLU 正常组 10 2.01 ± 0.37 1.22 ± 0.55 0.26 ± 0.08 1.44 ± 0.27 46.31 ± 14.93 7.12 ± 1.01 模型组 8 1.56 ± 0.22# 1.14 ± 0.27 0.21 ± 0.04 1.00 ± 0.22## 57.71 ± 20.51 8.04 ± 0.99 培哚普利低剂量组 8 1.58 ± 0.32 1.20 ± 0.21 0.22 ± 0.08 1.00 ± 0.26## 61.59 ± 12.65 7.76 ± 0.62 培哚普利高剂量组 8 1.60 ± 0.28 1.03 ± 0.39 0.26 ± 0.09 1.03 ± 0.20# 48.38 ± 15.50 7.36 ± 0.74 与正常组比较,#P < 0. 05, ##P < 0. 01。
下载: 导出CSV
表 3 培哚普利对大鼠肠道菌群Alpha多样性分析 ( $\bar x \pm s $)
Table 3. Analysis of Alpha diversity of rat intestinal flora by perindopril ( $\bar x \pm s $)
组别 n Shannon指数 Simpson指数 Ace指数 Chaol指数 Coverage 正常组 10 2.64 ± 0.80 0.21 ± 0.14 312.20 ± 98.20 310.03 ± 97.44 0.99 ± 0.00 模型组 8 3.43 ± 0.63 0.11 ± 0.06 377.88 ± 95.42 384.58 ± 97.28 0.99 ± 0.00 培哚普利低剂量组 8 3.51 ± 0.96 0.11 ± 0.11 421.71 ± 90.84# 433.69 ± 93.11# 0.99 ± 0.00 培哚普利高剂量组 8 3.06 ± 1.02 0.17 ± 0.19 344.09 ± 119.36 345.81 ± 126.86 0.99 ± 0.00 与正常组比较,#P < 0. 05。
下载: 导出CSV
表 4 各组大鼠粪便在门水平上的相对丰度 (%/ $\bar x \pm s $,正常组n = 10,其余组n = 8)
Table 4. Relative abundance of rat feces at phylum level in each group (%/ $\bar x \pm s $,normal group n = 10,other groups n = 8)
组别 厚壁菌门 拟杆菌门 脱硫杆菌门 放线菌门 弯曲杆菌门 正常组 94.07 ± 3.89* 1.61 ± 1.85 0.62 ± 0.59** 1.02 ± 0.70 0.03 ± 0.03 模型组 87.02 ± 6.39 5.81 ± 4.29 3.13 ± 1.71 0.55 ± 0.32 1.53 ± 3.09** 培哚普利低剂量组 67.79 ± 10.08##** 17.40 ± 13.90## 2.90 ± 2.86# 1.50 ± 1.40 0.76 ± 0.94# 培哚普利高剂量组 83.08 ± 7.50 8.11 ± 6.93 2.58 ± 2.72 2.04 ± 1.27** 0.39 ± 0.88 与正常组比较,#P < 0.05, ##P < 0.01;与模型组比较, *P < 0.05, **P < 0.01。
下载: 导出CSV
-
[1] Bl ü her M. Obesity:global epidemiology and pathogenesis[J]. Nat Rev Endocrino,2019,15(5):288-298. doi: 10.1038/s41574-019-0176-8 [2] World Health Organization. Obesity and overweight[EB/OL]. (2021-06-09)[2022-12-10]. https://www.who.int/en/news-room/fact-sheets/detail/obesity-and-overweight. [3] Zhang X,Zhang M,Zhao Z P,et al. Geographic variation in prevalence of adult obesity in China:Results from the 2013-2014 national chronic disease and risk factor surveillance[J]. Ann Intern Med,2019,172(4):291-293. [4] Wang Y,Zhao L,Gao L,et al. Health policy and public health implications of obesity in China[J]. Lancet Diabetes Endocrinol,2021,9(7):446-461. [5] Gasmi A,Mujawdiya P K,Pivina L,et al. Relationship between gut microbiota,gut hyperpermeability,and obesity[J]. Curr Med Chem,2020,28(28):827-839. [6] Canfora E E,Meex R C R,Venema K,et al. Gut microbial metabolites in obesity,NAFLD and T2DM[J]. Nature Reviews Endocrinology,2019,15(5):261-273. doi: 10.1038/s41574-019-0156-z [7] Li R,Huang X,Liang X,et al. Integrated omics analysis reveals the alteration of gut microbe -metabolites in obese adults[J]. Briefings in Bioinformatics,2020,22(3):1-16. [8] Kootte R S, Levin E, Salojarvi J, et al. Improvement of insulin sensitivity after lean donor feces in metabolic syndrome is driven by baseline intestinal microbiota composition[J]. Cell Metabolism, 2017, 26(4): 611-619. [9] 郭艳东. 云南高原偏寒地区汉族PPARγ2基因Pro12Ala、C161-T多态性与肥胖的关联研究[D]. 昆明: 昆明医科大学, 2020. [10] Jones B H,Standridgr M K,Moustaid N. Angiotensin II increases lipogenesis in 3T3-L1 and human adipose cells[J]. Endocrinolog,1997,138(4):1512-1519. doi: 10.1210/endo.138.4.5038 [11] Barton M, Carmona R, Morawietz H. Obesity is associated with tissue-specific activation of renal angiotensin-converting enzyme in vivo: Evidence for a regulatory role of endothelin[J]. Hypertension, 2000, 35(1): 329-336. [12] Rankinen T,Zuberi A,Chagnon Y C,et al. The human obesity gene map:The 2005 update[J]. Obesity,2006,14(4):529-644. doi: 10.1038/oby.2006.71 [13] Masuo K,Mikami H,Ogihara T,et al. Weight reduction and pharmacologic treatment in obese hypertensives[J]. Am J Hypertens,2001,14(6):530-538. doi: 10.1016/S0895-7061(00)01279-6 [14] Scholze J,Grimm E,Herrmann D,et al. Optimal treatment of obesity-related hypertension:the Hypertension-Obesity-Sibutramine (HOS) study[J]. Circulation,2007,115(15):1991-1998. doi: 10.1161/CIRCULATIONAHA.106.625400 [15] 吐克孜. 吾守尔,王茹,张姣姣,等. 樱桃李果实提取物对小鼠高脂膳食诱导肥胖的预防作用[J]. 中国食品学报,2021,21(6):144-149. [16] Katta N,Loethen T,Lavie C J,et al. Obesity and coronary heart disease:Epidemiology,pathology,and coronary artery imaging[J]. Current Problems in Cardiology,2020,46(3):1-43. [17] Ussar S,Griffin N W,Bezy O,et al. Interactions between gut microbiota,host genetics and diet modulate the predisposition to oObesity and metabolic syndrome[J]. Cell Metabolism,2015,22(3):516-530. doi: 10.1016/j.cmet.2015.07.007 [18] 叶晓琳,马亚楠,刘洋,等. 肠道菌群在肥胖发病中的作用[J]. 中国微生态学杂志,2020,32(11):1349-1353. [19] Chakraborti C K. New-found link between microbiota and obesity[J]. World J Gastrointest Pathophysiol,2015,6(4):110-119. doi: 10.4291/wjgp.v6.i4.110 [20] Jumpertz R,Le D S,Turnbaugh P J,et al. Energy-balance studies reveal associations between gut microbes,caloric load,and nutrient ab- sorption in humans[J]. Am J Clin Nutr,2011,94(1):58-65. doi: 10.3945/ajcn.110.010132 [21] Da Silva C C,Monteil M A,Davis E M. Overweight and obesity in chil- dren are associated with an abundance of firmicutes and reduction of Bifidobacterium in their gastrointestinal microbiota[J]. Child Obes,2020,16(3):204-210. doi: 10.1089/chi.2019.0280 [22] Ivarsson E,Roos S,Liu H Y,et al. Fermentable non-starch polysaccharides increases the abundance of Bacteroides-PrevotellaPorphyromonas in ileal microbial community of growing pigs[J]. Animal,2014,8(11):1777-1787. doi: 10.1017/S1751731114001827 [23] Chambers E S,Viardot A,Psichas A,et al. Effects of targeted delivery of propionate to the human colon on appetite regulation,body weight maintenance and adiposity in overweight adults[J]. Gut,2015,64(11):1-10. [24] Murugesan S,Nirmalkar K,Hoyovadillo C,et al. Gut microbiome production of short-chain fatty acids and obesity in children[J]. Eur J Clin Microbiol Infect Dis,2018,37(4):1-5. -


 点击查看大图
点击查看大图

计量
- 文章访问数: 3177
- HTML全文浏览量: 2241
- PDF下载量: 45
- 被引次数: 0
