

Study on Osteogenic Ability of Dental Pulp Stem Cells at Different Concentrations
-
摘要:
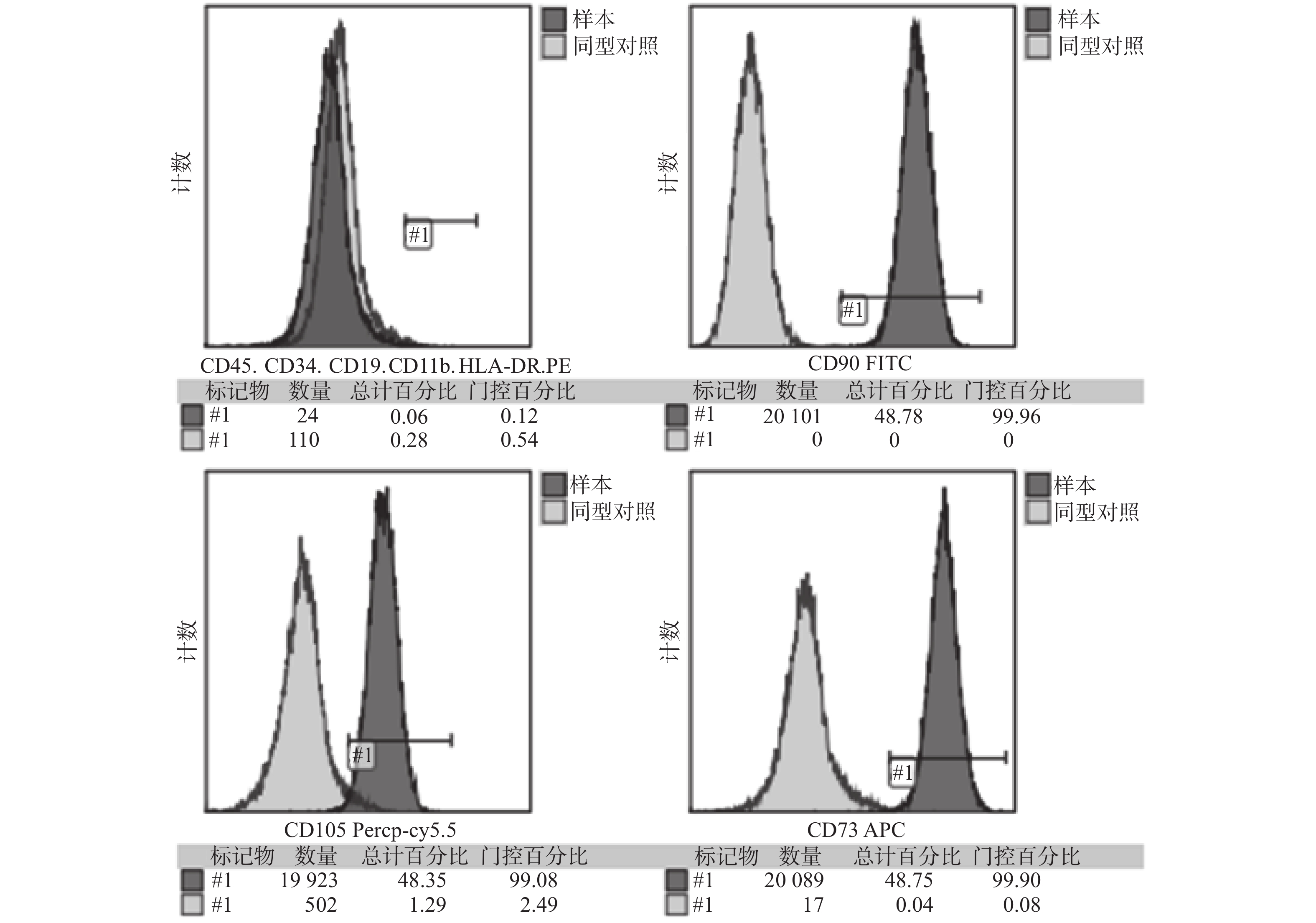
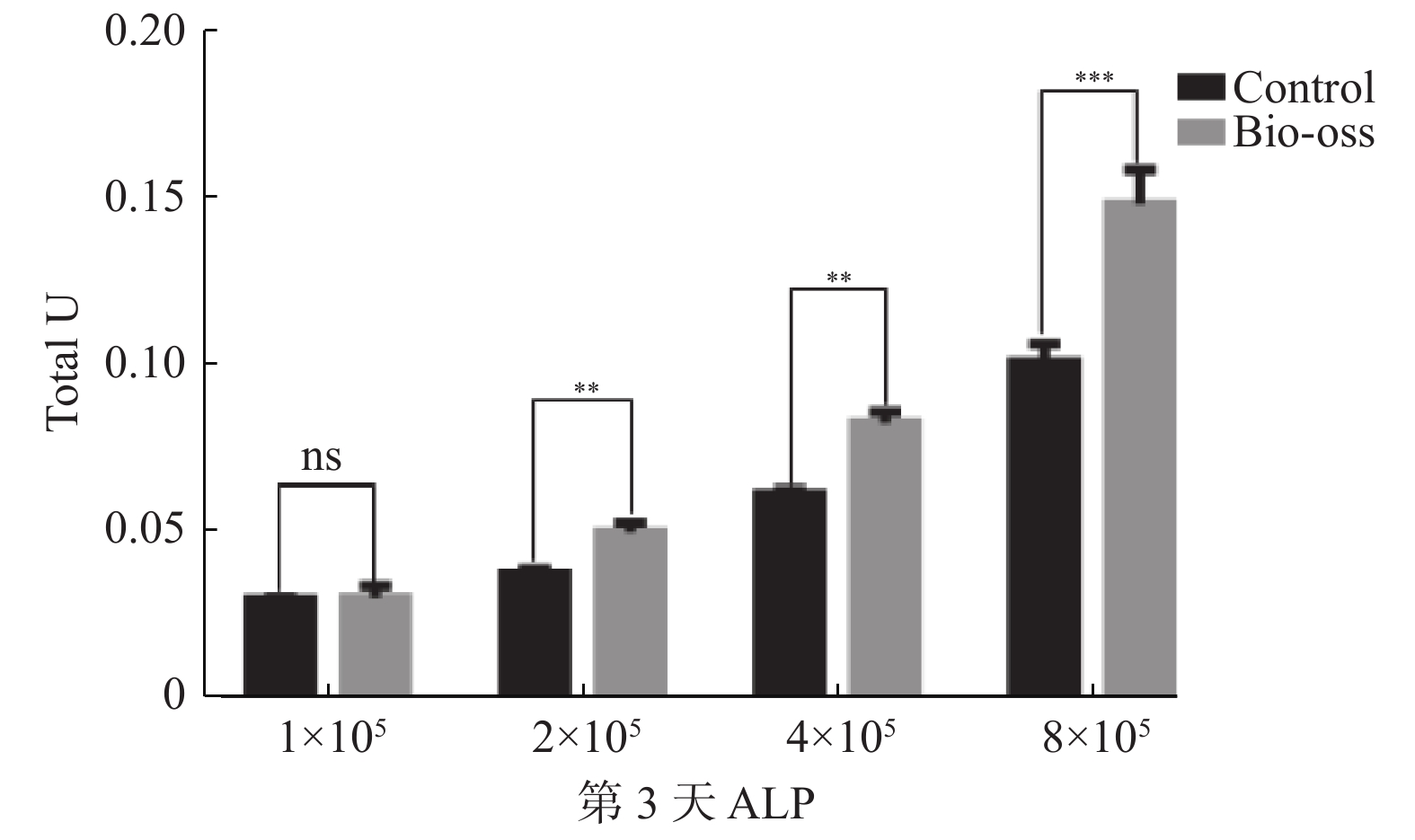
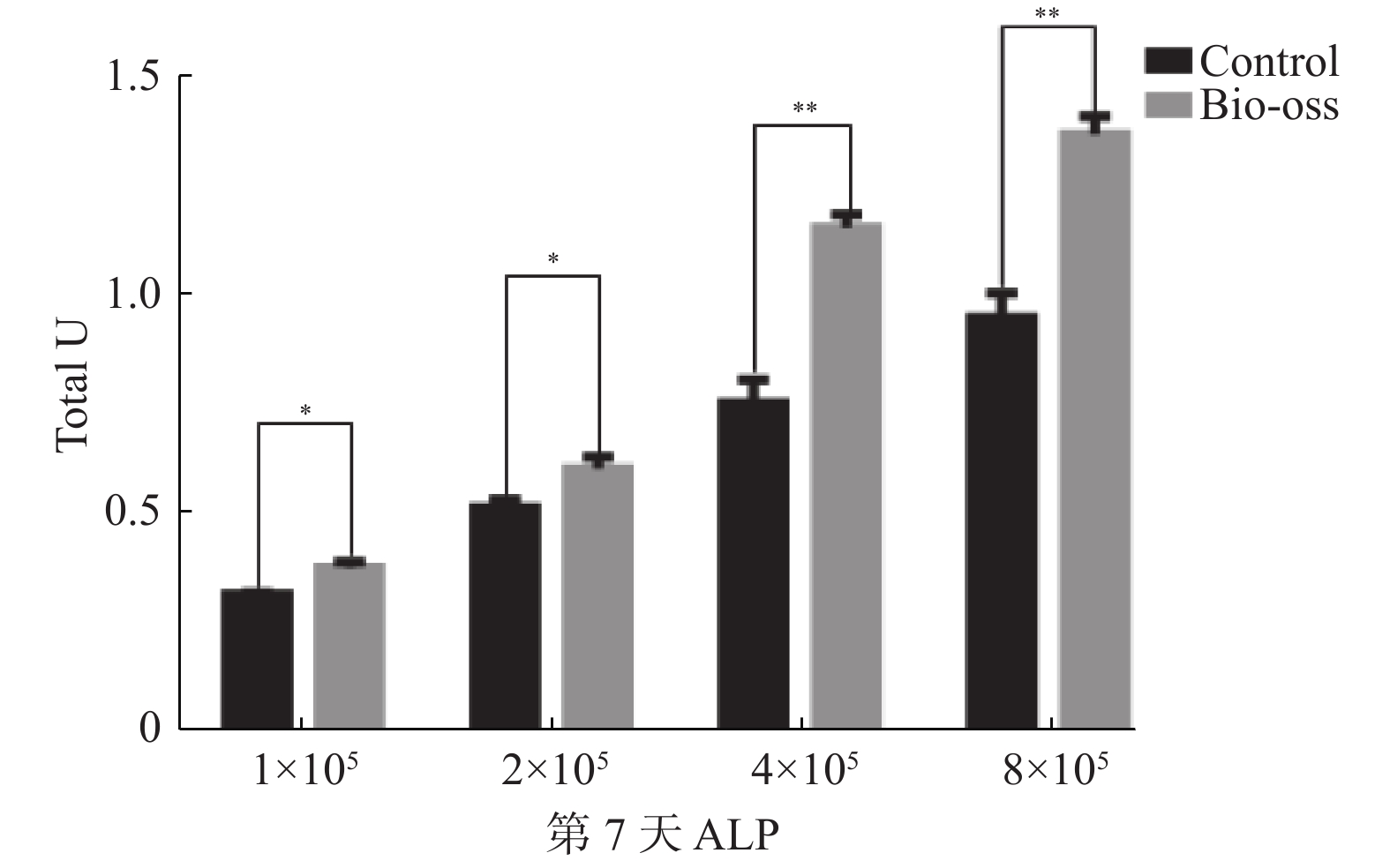
目的 探究不同浓度的牙髓干细胞(dental pulp stem cell,DPSC)在定量Bio-Oss骨粉中最佳的成骨比例。 方法 收集牙髓组织、组织块贴壁法分离培养原代细胞,流式细胞术鉴定DPSC表面标志物,诱导成骨及成脂鉴定DPSC多向分化潜能;GFP慢病毒转染DPSC,嘌呤霉素筛选GFP-DPSC;设置1×105、2×105、4×105、8×1054组细胞数接种于0.05 g骨粉并成骨培养,对照组不加骨粉,第3天和第7天进行碱性磷酸酶活性测定;培养第7天在扫描电镜下观察细胞在骨粉内的形态变化。 结果 原代分离培养成功的DPSC符合间充质干细胞来源,相关表面标记物高表达,可向成脂和成骨分化;GFP成功转染DPSC,第3天和第7天进行碱性磷酸酶活性检测,实验组均高于对照组,差异具有统计学意义(P < 0.05);扫描电镜发现DPSC在骨粉表面爬行,部分细胞伸出伪足,4×10 5和8×105 2组细胞较为密集。 结论 4×105~8×105个DPSC与0.05 g Bio-Oss骨粉复合培养成骨效果较好,可以缩短成骨周期,细胞若接种过多,成骨效果反而抑制,造成干细胞的浪费。 Abstract:Objective To explore the optimal osteogenic ratio of Dental pulp stem cells (DPSC) with different concentrations in quantitative Bio-Oss bone powder. Methods The primary cells were isolated and cultured by adherent method. The surface markers of DPSC were identified by flow cytometry, and the multidirectional differentiation potential of DPSC was identified by osteogenesis and adipogenesis. DPSC was transfected with GFP lentivirus, and GFP-DPSC was screened with puromycin. The cells in groups 1×105, 2×105, 4×105 and 8×105 were inoculated with 0.05 g bone powder and cultured for osteogenesis, while the control group did not add bone powder. Alkaline phosphatase activity was measured on day 3 and day 7. On the 7th day of culture, the morphological changes of the cells in bone powder were observed under scanning electron microscope. Results The DPSC was derived from mesenchymal stem cells with high expression of related surface markers and could differentiate into adipogenic and osteogenic cells. GFP was successfully transfected into DPSC, and alkaline phosphatase activity was detected on the 3rd and 7th day, and the experimental group was higher than the control group, and the difference was statistically significant (P < 0.05). Scanning electron microscopy showed that DPSC crawled on the surface of bone powder, and some cells extended pseudopodia, and the cells of 4×10 5 and 8×105 groups were more dense. Conclusions The combined culture of 4×105-8×105 DPSC and 0.05 g Bio-Oss bone powder has a good osteogenic effect, which can shorten the osteogenic cycle. If the cells are inoculated too much, the osteogenic effect is inhibited, resulting in the waste of stem cells. -

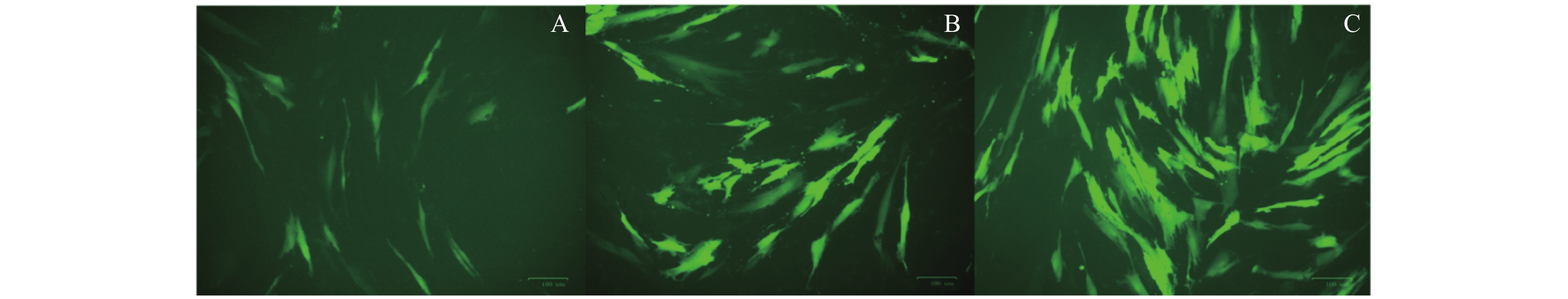
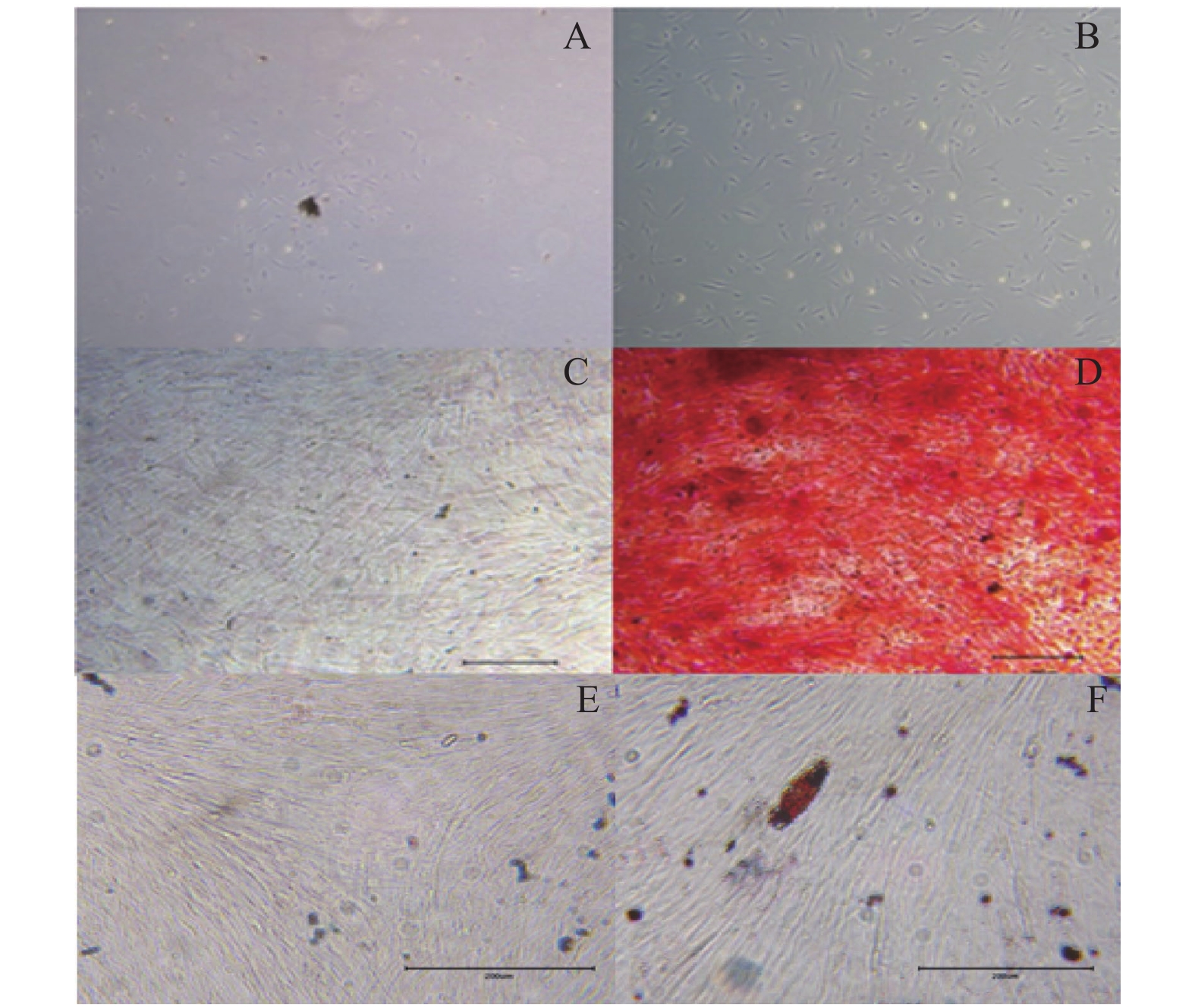
图 1 DPSC的培养及多向分化能力鉴定
A:DPSC原代细胞(×50);B:扩大培养第三代细胞(×50);C:DPSC成骨对照组组染色所见(×100);D:DPSC成骨诱导组染色所见(×100);E:DPSC成脂对照组染色所见(×200);F:DPSC成脂诱导组染色所见(×200)。
Figure 1. Culture of DPSC and appraisal of multi -directional differentiation ability

图 3 GFP转染DPSC不同时间荧光表达(×100)
A:36 h GFP转染DPSC的荧光表达;B:72 h GFP转染DPSC的荧光表达;C:96 h GFP转染DPSC的荧光表达。
Figure 3. Fluorescence expression of GFP transfected DPSC at different time
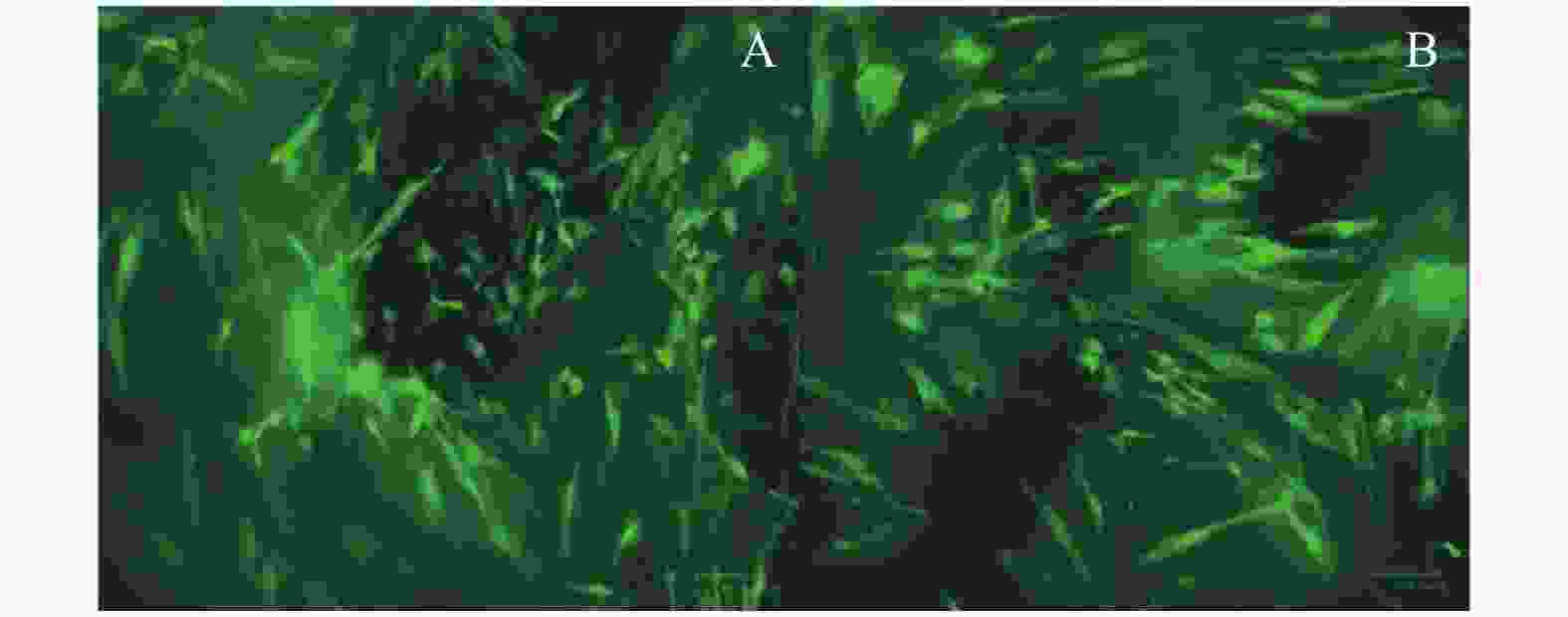
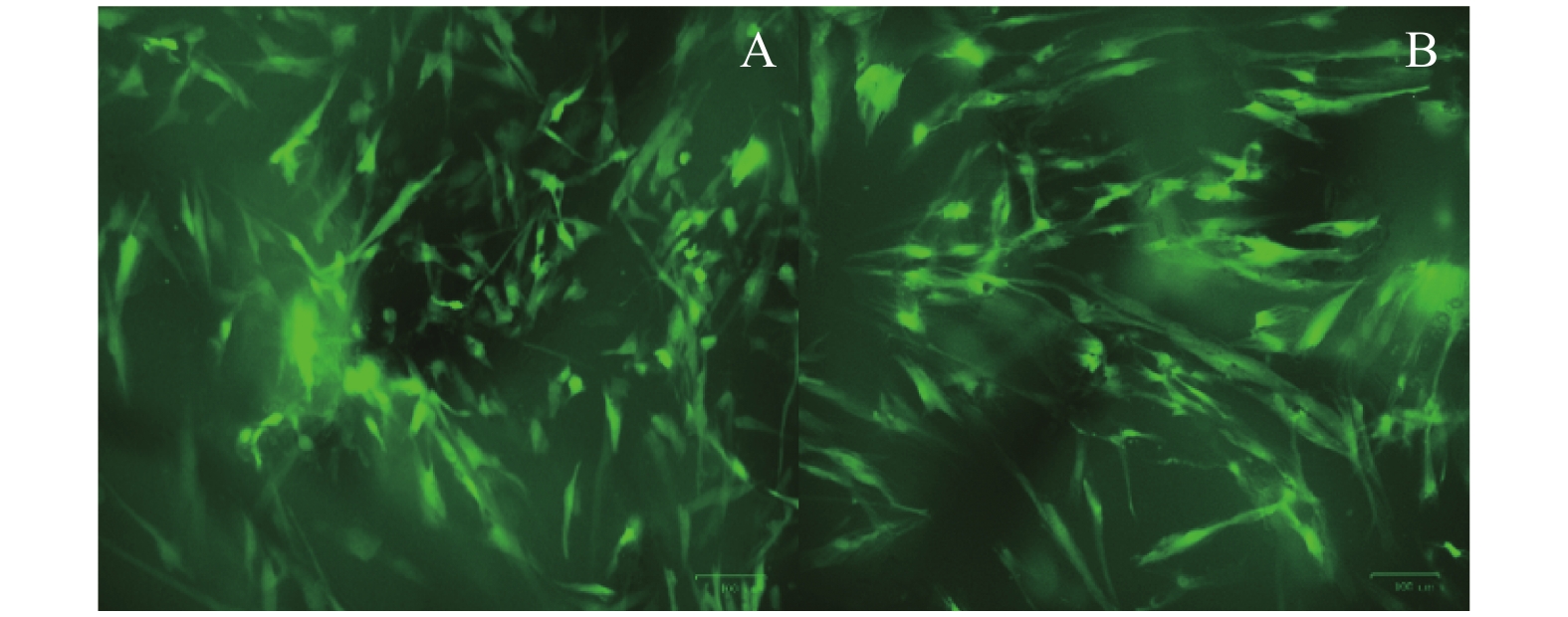
图 4 GFP-DPSC在Bio-Oss骨粉中的生长(×100)
A:第3天时GFP-DPSC在Bio-Oss骨粉中的状态;B:第14天时GFP-DPSC在Bio-Oss骨粉中的状态。
Figure 4. Growth of GFP-DPSC in Bio-Oss bone powder

图 7 1×105组同一视野下SEM图像
A:100 μm ;B:50 μm。
Figure 7. SEM images of 1×105 groups in the same field of view

图 8 2×105组同一视野下SEM图像
A:100 μm ;B:50 μm。
Figure 8. SEM images of 2×105 groups in the same field of view

图 9 4×105组同一视野下SEM图像
A:100 μm ;B:50 μm。
Figure 9. SEM images of 4×105 groups in the same field of view
-
[1] Le Thieu M K,Homayouni A,Haeren L R,et al. Impact of simultaneous placement of implant and block bone graft substitute: an in vivo peri-implant defect model[J]. Biomater Res,2021,25(1):43-52. doi: 10.1186/s40824-021-00245-3 [2] Shiu S T,Lee W F,Chen S M,et al. Effect of different bone grafting materials and mesenchymal stem cells on bone regeneration: A micro-computed tomography and histomorphometric study in a rabbit calvarial defect model[J]. Int J Mol Sci,2021,22(15):8101-8115. doi: 10.3390/ijms22158101 [3] Shi X,Mao J,Liu Y. Pulp stem cells derived from human permanent and deciduous teeth: Biological characteristics and therapeutic applications[J]. Stem Cells Transl Med,2020,9(4):445-464. doi: 10.1002/sctm.19-0398 [4] Tsutsui T W. Dental pulp stem cells: advances to applications[J]. Stem Cells Cloning,2020,13:33-42. [5] Gendviliene I,Simoliunas E,Alksne M,et al. Effect of extracellular matrix and dental pulp stem cells on bone regeneration with 3D printed PLA/HA composite scaffolds[J]. Eur Cell Mater,2021,41:204-215. doi: 10.22203/eCM.v041a15 [6] Tanikawa D Y S,Pinheiro C C G,Almeida M C A,et al. Deciduous Dental Pulp Stem Cells for Maxillary Alveolar Reconstruction in Cleft Lip and Palate Patients[J]. Stem Cells Int,2020,2020:6234167. [7] Gil L F,Nayak V V,Benalcazar Jalkh E B,et al. Laddec(R) versus Bio-Oss(R): The effect on the healing of critical-sized defect - Calvaria rabbit model[J]. J Biomed Mater Res B Appl Biomater,2022,110(12):2744-2750. doi: 10.1002/jbm.b.35125 [8] Li Y,Zhou W,Li P,et al. Comparison of the osteogenic effectiveness of an autogenous demineralised dentin matrix and Bio-Oss(R) in bone augmentation: a systematic review and meta-analysis[J]. Br J Oral Maxillofac Surg,2022,60(7):868-876. doi: 10.1016/j.bjoms.2022.03.009 [9] Dai Y,Xu J,Han X H,et al. Clinical efficacy of mineralized collagen (MC) versus anorganic bovine bone (Bio-Oss) for immediate implant placement in esthetic area: a single-center retrospective study[J]. BMC Oral Health,2021,21(1):390-398. doi: 10.1186/s12903-021-01752-4 [10] Shamsoddin E,Houshmand B,Golabgiran M. Biomaterial selection for bone augmentation in implant dentistry: A systematic review[J]. J Adv Pharm Technol Res,2019,10(2):46-50. doi: 10.4103/japtr.JAPTR_327_18 [11] RAPONE B,INCHINGOLO A D,TRASARTI S,et al. Long-Term Outcomes of Implants Placed in Maxillary Sinus Floor Augmentation with Porous Fluorohydroxyapatite (Algipore((R)) FRIOS((R))) in Comparison with Anorganic Bovine Bone (Bio-Oss((R))) and Platelet Rich Plasma (PRP): A Retrospective Study[J]. J Clin Med,2022,11(9):2491-2503. doi: 10.3390/jcm11092491 [12] Iaquinta M R,Martini F,D'agostino A,et al. Stem cell fate and immunomodulation promote bone regeneration via composite bio-oss((R))/avitene(TM) biomaterial[J]. Front Bioeng Biotechnol,2022,10:873814. doi: 10.3389/fbioe.2022.873814 [13] Kosinski M,Figiel-dabrowska A,Lech W,et al. Bone defect repair using a bone substitute supported by mesenchymal stem cells derived from the umbilical cord[J]. Stem Cells Int,2020,2020:1321283. [14] Jiang Y H,Shang Y,Zou D H,et al. [Effect of rat allogeneic BMSCs-Bio-Oss-bFGF compound on tooth extraction healing: a micro-CT study][J]. Shanghai Kou Qiang Yi Xue,2022,31(1):38-43. [15] Zhou Q,Yu B H,Liu W C,et al. BM-MSCs and bio-oss complexes enhanced new bone formation during maxillary sinus floor augmentation by promoting differentiation of BM-MSCs[J]. In Vitro Cell Dev Biol Anim,2016,52(7):757-771. doi: 10.1007/s11626-015-9995-7 [16] Xu X,Fang K,Wang L,et al. Local application of semaphorin 3a combined with adipose-derived stem cell sheet and anorganic bovine bone granules enhances bone regeneration in type 2 diabetes mellitus rats[J]. Stem Cells Int,2019,2019:2506463. [17] Chen Y,Huang H,Li G,et al. Dental-derived mesenchymal stem cell sheets: a prospective tissue engineering for regenerative medicine[J]. Stem Cell Res Ther,2022,13(1):38-52. doi: 10.1186/s13287-022-02716-3 [18] Alksne M,Kalvaityte M,Simoliunas E,et al. Dental pulp stem cell-derived extracellular matrix: autologous tool boosting bone regeneration[J]. Cytotherapy,2022,24(6):597-607. doi: 10.1016/j.jcyt.2022.02.002 [19] Davies O G,Cooper P R,Shelton R M,et al. A comparison of the in vitro mineralisation and dentinogenic potential of mesenchymal stem cells derived from adipose tissue,bone marrow and dental pulp[J]. J Bone Miner Metab,2015,33(4):371-382. doi: 10.1007/s00774-014-0601-y [20] Lee Y C,Chan Y H,Hsieh S C,et al. Comparing the osteogenic potentials and bone regeneration capacities of bone marrow and dental pulp mesenchymal stem cells in a rabbit calvarial bone defect model[J]. Int J Mol Sci,2019,20(20):5015-5024. doi: 10.3390/ijms20205015 [21] Lorusso F,Inchingolo F,Dipalma G,et al. Synthetic scaffold/dental pulp stem cell (DPSC) tissue engineering constructs for bone defect treatment: an animal studies literature review[J]. Int J Mol Sci,2020,21(24):9765-9784. doi: 10.3390/ijms21249765 [22] Wang F,Li Q,Wang Z. A comparative study of the effect of Bio-Oss((R)) in combination with concentrated growth factors or bone marrow-derived mesenchymal stem cells in canine sinus grafting[J]. J Oral Pathol Med,2017,46(7):528-536. doi: 10.1111/jop.12507 [23] Qin R,Cui Z,Zhou H,et al. Effect of lentivirus-mediated BMP2 from autologous tooth on the proliferative and osteogenic capacity of human periodontal ligament cells[J]. J Periodontal Res,2022,57(4):869-879. doi: 10.1111/jre.13025 [24] Zhou C,Zhang D,Zou J,et al. Substrate Compliance Directs the Osteogenic Lineages of Stem Cells from the Human Apical Papilla via the Processes of Mechanosensing and Mechanotransduction[J]. ACS Appl Mater Interfaces,2019,11(29):26448-26459. doi: 10.1021/acsami.9b07147 [25] Shahabipour F,Oskuee R K,Shokrgozar M A,et al. CRISPR/Cas9 mediated GFP-human dentin matrix protein 1 (DMP1) promoter knock-in at the ROSA26 locus in mesenchymal stem cell for monitoring osteoblast differentiation[J]. J Gene Med,2020,22(12):e3288. -

 下载:
下载:




 点击查看大图
点击查看大图
计量
- 文章访问数: 3354
- HTML全文浏览量: 2161
- PDF下载量: 24
- 被引次数: 0
