

Comparison of Two Induction Methods of Temporo-mandibular Joint Osteoarthritis in Rat Models
-
摘要:
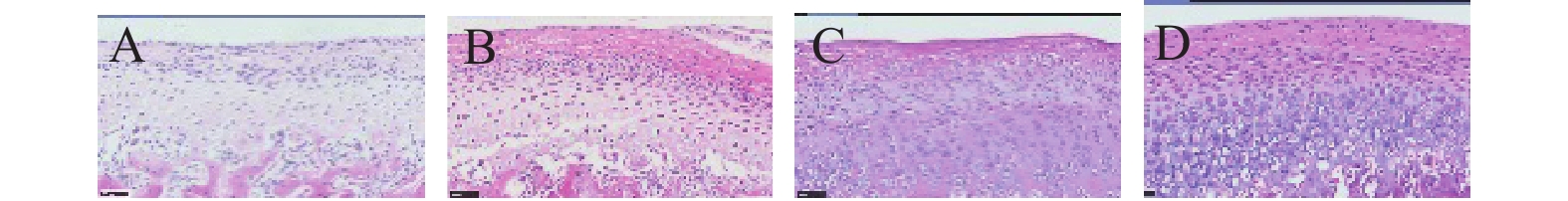
目的 通过被动张口和咬合紊乱2种方法建立大鼠颞下颌关节骨关节炎的模型,观察软骨和软骨下骨的病理变化来比较2种建模方法并评价其实用性。 方法 21只8周龄雄性SD大鼠随机分为4组,对照组(Control组)3只大鼠常规饲养。剩余18只大鼠随机分成3组:被动张口组(OP组),每天保持张口度20 mm,持续1 min;咬合紊乱组(OD组),0.25 mm的结扎丝包绕第一磨牙,结扎在第一磨牙的面;咬合紊乱对照组(ODS组)采用同样的手术方法,结扎结在第一磨牙的近中。4周后处死所有大鼠,解剖下颌骨后,通过Micro-CT,番红-O-固绿染色,HE染色来评估颞下颌关节的变化。 结果 OD组和OP组均出现了颞下颌关节骨关节炎样组织学改变,Micro CT分析都表现出骨体积分数的下降,骨体积比和骨小梁厚度在OD组与OP组的差异无统计学意义(P > 0.05)。OD组和OP组的改良Mankin评分均增高,但差异无统计学意义(P > 0.05),且OP组中有2个样本未出现颞下颌关节骨关节炎样组织学改变。 结论 咬合紊乱为较为稳定的大鼠颞下颌关节骨关节炎的建模方法,被动张口能造成大鼠颞下颌关节骨关节炎的组织病理学表现,但不稳定且效率较低。 Abstract:Objective To establish TMJ-OA rat models by occlusal disorder and forced open mouth, compare two TMJ-OA induction methods and assess their applicability by pathological changes in the cartilage, subchondral bone. Methods Twenty-one 8-week-old Sprague-Dawley (SD) male rats were divided into four groups, namely control group, and three rats were conventional feeding. eighteen SD rats were randomly allocated to three groups using the randomization table; each group had the same number of SD rats. The Forced open mouth (OP) , steady mouth opening was imposed to 20 mm daily (1 minute/day for 16 days). the occlusal disorder group (OD), an orthodontic ligation silk (0.25 mm diameter) revolved around the first molar, and a ligation knot was created on the first molar of the maxillary. The control group of occlusal disorder (ODS) use the same method, a ligation knot in the mesial of the first molar. Rat models were employed and followed for 4 weeks after experimental procedures, and TMJ joints in each group were harvested. The TMJ changes were evaluated by micro-CT, HE staining, and Safranin-O/Fast green staining. Results We found apparent histological phenotypes of TMJ-OA in the OP and OD groups, and observed a pronounced drop in bone volume fraction , BV/TV and Tb.Th between OD and OP groups had no significant difference (P > 0.05), and a substantial increase in the modified Mankin score was found, there was no significant difference in the two groups (P > 0.05), but two samples of OP group didn’t present apparent histological phenotypes of TMJ-OA. Conclusions Osteoarthritis-like pathological changes in the cartilage and subchondral bone can be observed in both of the two methods. The occlusal disorder is a stable TMJ-OA model, forced open mouth caused osteoarthritis-like changes, but it’s unstable and inefficient. -
Key words:
- Temporomandibular disorders /
- Osteoarthritis /
- Temporomandibular joint /
- Rat models /
- Occlusal disorder
-

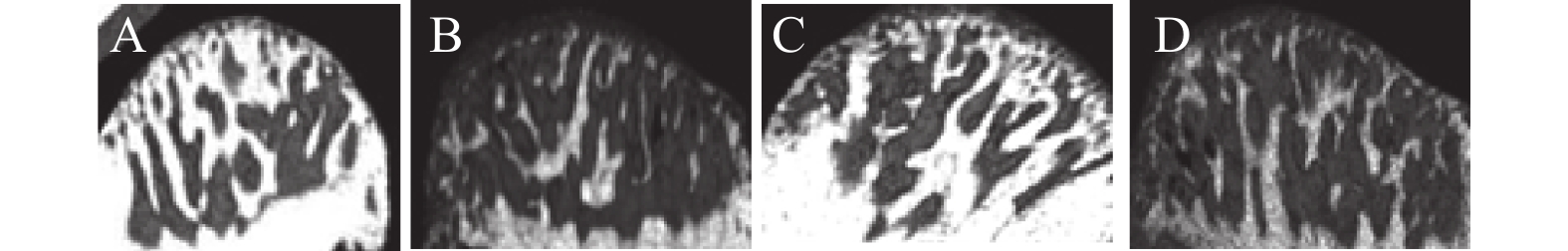
图 3 骨体积比 (%,BV/TV) 以及骨小梁厚度 Tb.Th(mm)对比
*P < 0.05;**P < 0.01;ns:P > 0.05。
Figure 3. Comparison of BV/TV(%) and Tb.Th(mm)

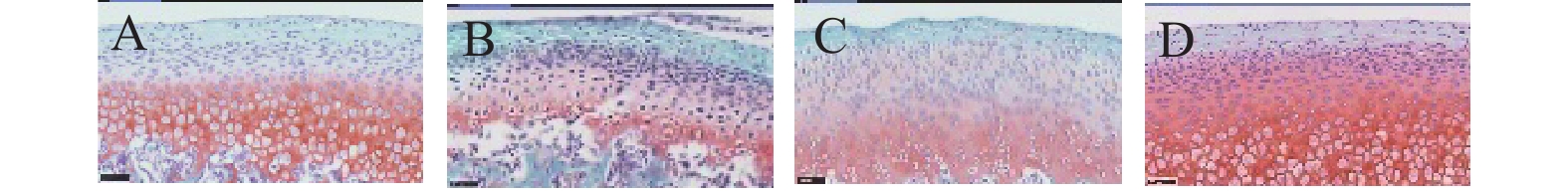
图 5 颞下颌关节番红-O-固绿染色(40×)
A:control组;B:OP组;C:ODS组;D:OD组。
Figure 5. TMJ Safranin-O/Fast green staining (40×)

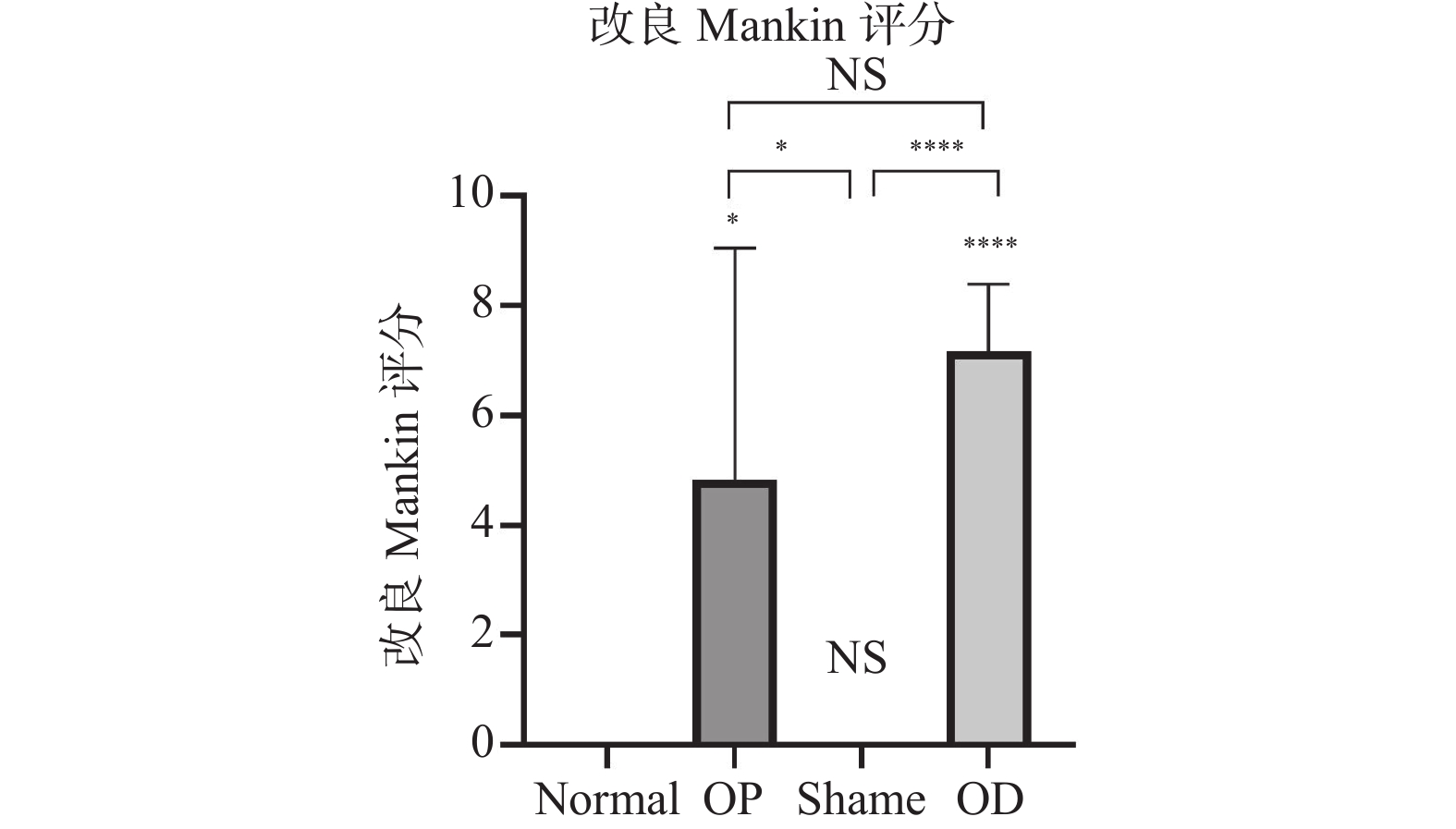
图 6 改良Mankin评分比较
ns:P > 0.05;*P < 0.05;****P < 0.001。
Figure 6. Comparison of modified mankin score
表 1 关节软骨改良Mankin评分标准
Table 1. Modified mankin score of articular cartilage
类别 分数 结构 正常 0 表层软骨破坏 1 血管翳及表层破坏 2 浅层裂隙形成达移行层 3 裂隙局限性深达骨质辐射层 4 深达骨质钙化层 5 全层软骨缺损 6 番红-O-固绿染色 正常 0 轻度失染 1 中度失染 2 重度失染 3 完全失染 4 细胞 正常 0 细胞过多,紊乱 1 细胞成簇 2 细胞少 3 潮线完整性 完整 0 血管通过不完整 1 总分 0~14分 评分结果是通过对每个部分进行2次以上评分的平均值来确定。  下载: 导出CSV
下载: 导出CSV
-
[1] Zhao Y, An Y, Zhou L, et al. Animal models of temporomandibular joint osteoarthritis: Classification and selection[J]. Front Physiol 2022, 13: 859517. doi: 10.3389/fphys.2022.859517. PMID: 35574432. [2] Yuan W,Wu Y,Zhou X,et al. Comparison and applicability of three induction methods of temporomandibular joint osteoarthritis in murine models[J]. J Oral Rehabil,2022,49(4):430-441. doi: 10.1111/joor.13300.Epub2021Dec30 [3] de Souza R F,Lovato da Silva C H,Nasser M,et al. Interventions for the management of temporomandibular joint osteoarthritis[J]. Cochrane Database Syst Rev,2012,4:CD007261. doi: 10.1002/14651858.CD007261.pub2 [4] 杨文英,张文丽,罗应伟. 颞下颌关节骨关节炎动物模型的研究进展[J]. 国际口腔医学杂志,2015,42(6):677-680. [5] Almarza A J,Brown B N,Arzi B,et al. Preclinical animal models for temporomandibular joint tissue engineering[J]. Tissue Eng Part B Rev,2018,24(3):171-178. doi: 10.1089/ten.TEB.2017.0341.Epub2018Jan2 [6] Mankin H J,Dorfman H,Lippiello L,et al. Biochemical and metabolic abnormalities in articular cartilage from osteo-arthritic human hips. II. Correlation of morphology with biochemical and metabolic data[J]. J Bone Joint Surg Am,1971,53(3):523-537. doi: 10.2106/00004623-197153030-00009 [7] Mankin H J,Lippiello L. Biochemical and metabolic abnormalities in articular cartilage from osteo-arthritic human hips[J]. J Bone Joint Surg Am,1970,52(3):424-434. doi: 10.2106/00004623-197052030-00002 [8] Li B,Guan G,Li M,et al. Pathological mechanism of chondrocytes and the surrounding environment during osteoarthritis of temporomandibular joint[J]. Cell Mol Med,2021,25(11):4902-4911. doi: 10.1111/jcmm.16514 [9] Wang Q P,Yang L,Li X P,et al. Effects of 17β-estradiol on adiponectin regulation of the expression of osteoprotegerin and receptor activator of nuclear factor-κB ligand[J]. Bone,2012,51(3):515-523. doi: 10.1016/j.bone.2012.05.011.Epub2012May23 [10] Zhang J, Zhang S, Qi W J, et al. Mechanism and potential contributing factors to temporomandibular joint osteoarthritis[J]. Oral Dis, 2021. doi: 10.1111/odi.14061. Epub ahead of print. PMID: 34716969. [11] Liu Y D,Liao L F,Zhang H Y,et al. Reducing dietary loading decreases mouse temporomandibular joint degradation induced by anterior crossbite prosthesis[J]. Osteoarthritis Cartilage,2014,22(2):302-312. doi: 10.1016/j.joca.2013.11.014.Epub2013Dec5 [12] Zhang M,Wang H,Zhang J,et al. Unilateral anterior crossbite induces aberrant mineral deposition in degenerative temporomandibular cartilage in rats[J]. Osteoarthritis Cartilage,2016,24(5):921-931. doi: 10.1016/j.joca.2015.12.009.Epub2015Dec31 [13] 党薇,于世宾,何惠明,等. 不同开口度对大鼠咬肌及颞下颌关节的影响[J]. 牙体牙髓牙周病学杂志,2013,23(2):100-103,106. doi: 10.15956/j.cnki.chin.j.conserv.dent.2013.02.008 [14] 孙莲,张卫兵,王林. 开诱导大鼠颞下颌关节骨关节炎样退变的实验性研究[J]. 口腔医学,2016,36(5):390-393,410. doi: 10.13--591/j.cnki.kqyx.2016.05.002 [15] Muto T,Kawakami J,Kanazawa M,et al. Histologic study of synovitis induced by trauma to the rat temporomandibular joint[J]. Oral Surg Oral Med Oral Pathol Oral Radiol Endod,1998,86(5):534-540. doi: 10.1016/s1079-2104(98)90342-0 [16] Sun L,Wang M,He J,et al. Experimentally created nonbalanced occlusion effects on the thickness of the temporomandibular joint disc in rats[J]. Angle Orthod,2009,79(1):51-53. doi: 10.2319/091907-450.1 -


 点击查看大图
点击查看大图

计量
- 文章访问数: 5004
- HTML全文浏览量: 2261
- PDF下载量: 90
- 被引次数: 0
