

Mechanisms of Ginsenoside Rg1 in Protecting Cardiomyocytes by Sestrin2
-
摘要:
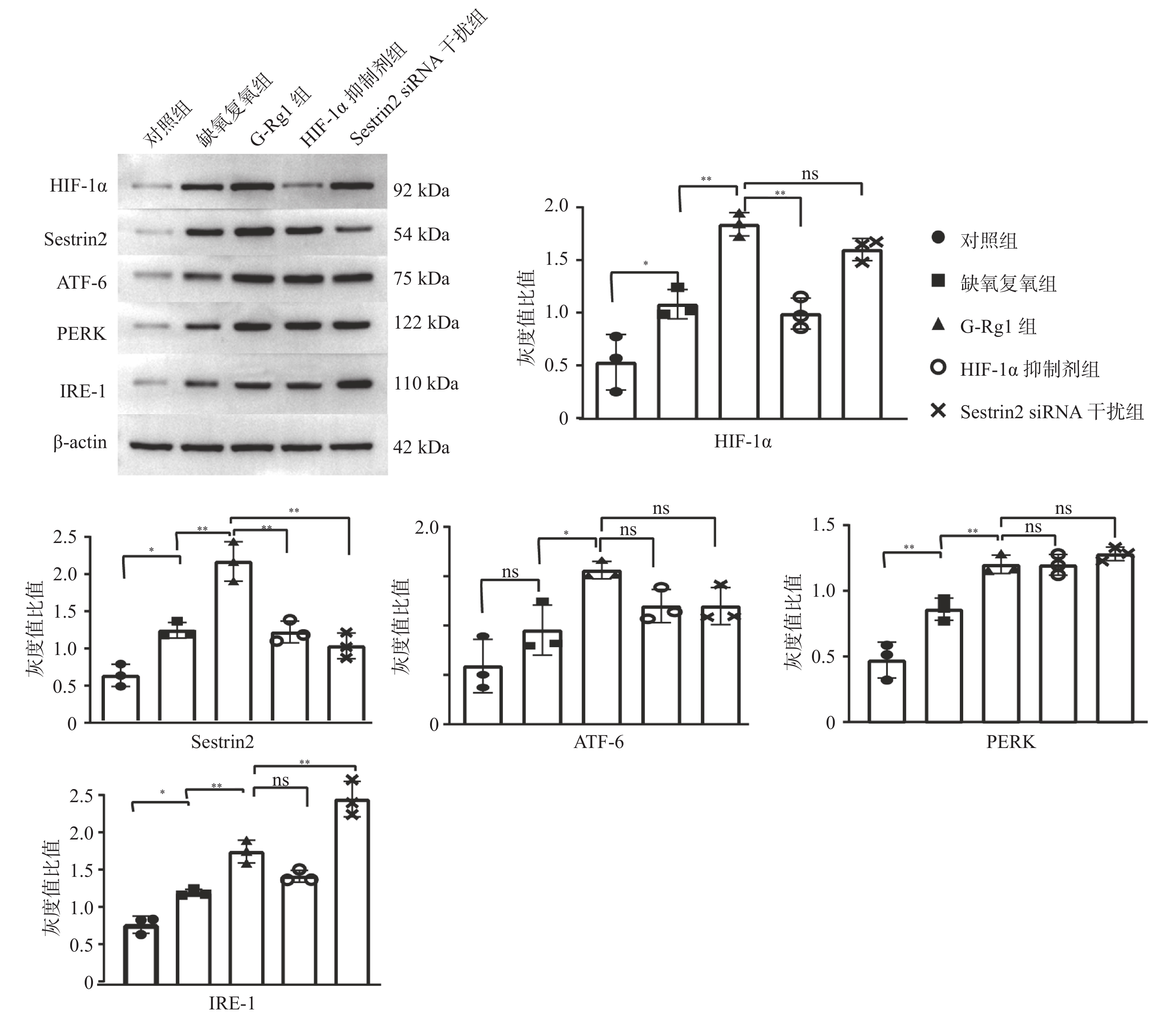
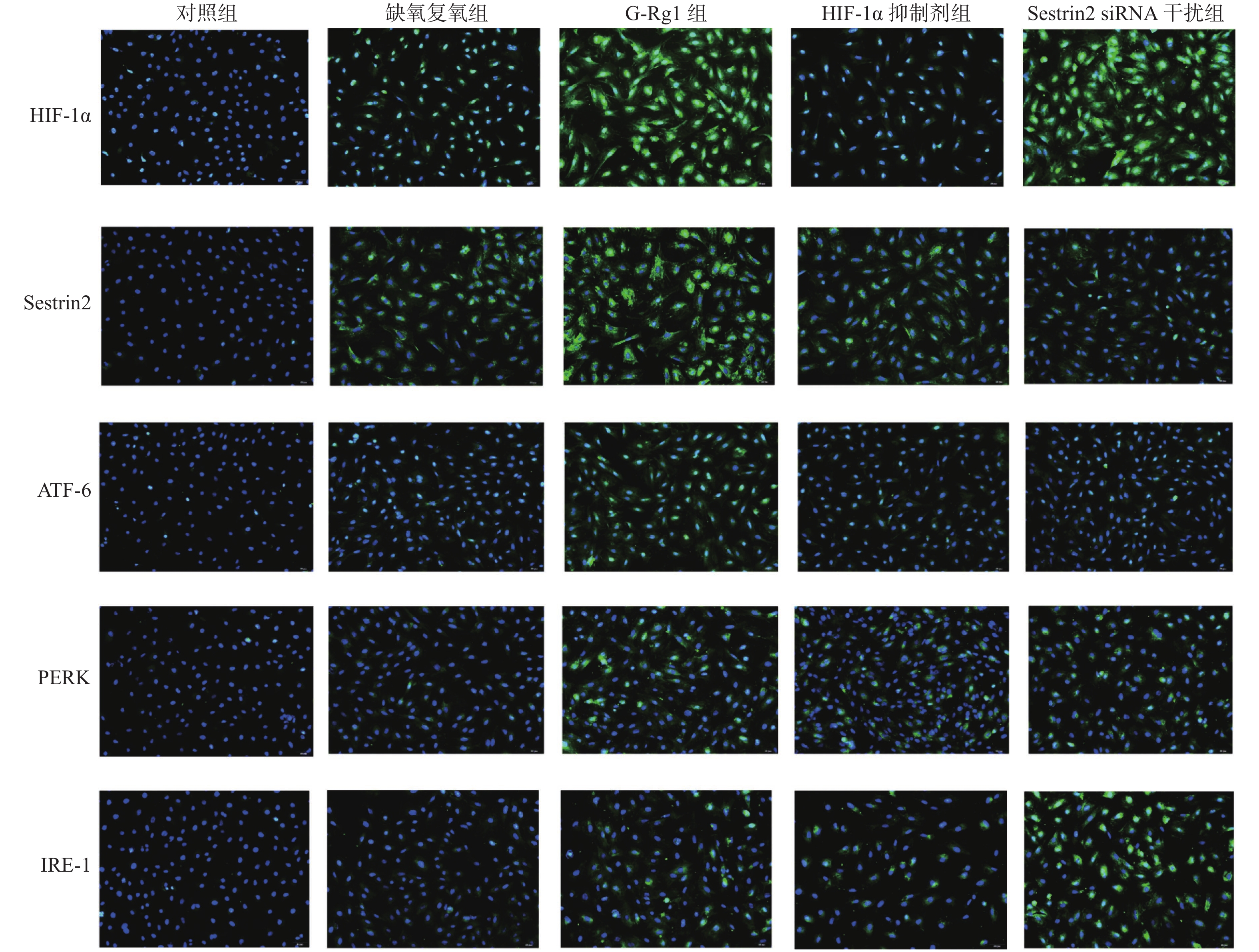
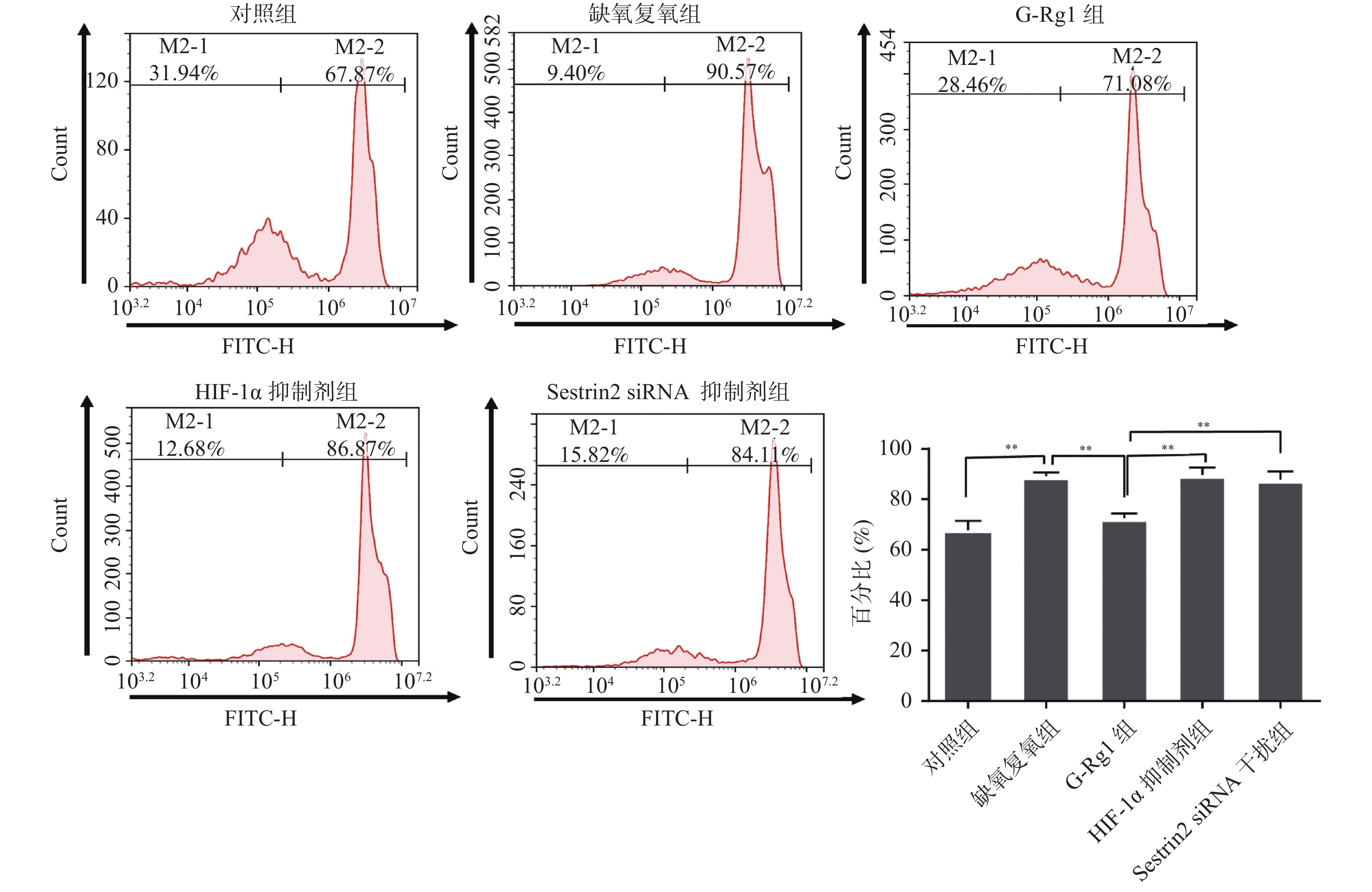
目的 研究人参皂苷Rg1(G-Rg1)对抗心肌缺血再灌注损伤中Sestrin2的作用及相关机制。 方法 体外培养大鼠 H9c2 心肌细胞,按实验要求随机分5组,分别为对照组,缺氧复氧组,G-Rg1组,缺氧诱导因子 -1α(Hypoxia Inducible Factor-1,HIF-1,HIF-1α) 抑制剂组,Sestrin2 siRNA干扰组。流式细胞术检测各组心肌细胞内活性氧簇(reactive oxygen species,ROS)含量。蛋白质免疫印迹法检测HIF-1α、Sestrin2、蛋白激酶R样内质网激酶(protein kinase R-like ER kinase,PERK)、活化转录因子6(the activating transcription factor 6,ATF-6)和跨膜蛋白激酶1(inositol requiring enzyme 1,IRE-1)蛋白表达。免疫荧光法检测Sestrin2、HIF-1α、IRE-1蛋白表达。 结果 与对照组相比,缺氧复氧组细胞内ROS水平、HIF-1α、Sestrin2、ATF-6、PERK、IRE-1蛋白表达水平明显升高(P < 0.05);与缺氧复氧组相比,G-Rg1组细胞内ROS水平明显降低(P < 0.05),HIF-1α、Sestrin2、ATF-6、PERK、IRE-1蛋白表达水平明显升高(P < 0.05);与G-Rg1组比较,HIF-1α抑制剂组心肌细胞内ROS水平升高,Sestrin2蛋白表达明显下调(P < 0.05);与G-Rg1组比较,siRNA干扰组心肌细胞内ROS水平升高,IRE-1表达明显升高(P < 0.05)。 结论 G-Rg1通过上调Sestrin2表达,保护缺氧复氧心肌细胞,并维持内质网应激平衡稳态。 Abstract:Objective To explore the role and mechanisms of Sestrin2 in ameliorating hypoxia-reoxygenation injury of rat’s cardiomyocytes by ginsenoside-rg1 (G-Rg1). Methods Rat H9c2 cardiomyocytes cultured in vitro were randomly divided into 5 groups according to the experimental requirements: control group, Hypoxia reoxygenation group, G-Rg1 group, hypoxia Inducible Factor-1 (HIF-1, HIF-1α) inhibitor group, and Sestrin2 siRNA interference group. Flow cytometry was used to detect the expression levels of ROS in cardiomyocytes. Western blot was used to detect the protein expression of HIF-1α, Sestrin2, ATF-6, PERK, IRE-1 in cardiomyocytes. Immunofluorescence was used to test the protein expression of Sestrin2, HIF-1α, IRE-1. Results Compared with control group, the level of ROS and protein expression of HIF-1α、Sestrin2、ATF-6、PERK、IRE-1 of hypoxiareoxygenation group were significantly increased (P < 0.05) . Compared with the hypoxiareoxygenation group , the ROS level in G-Rg1 group were significantly reduced and protein expression of HIF-1α、Sestrin2、ATF-6、PERK、IRE-1 in G-Rg1 group were significantly increased (P < 0.05) . Compared with G-Rg1 group, the level of ROS were significantly increased and the protein level of Sestrin2 were significantly reduced, in HIF-1α inhibition group (P < 0.05). Compared with G-Rg1 group, the level of ROS were significantly increased and the protein level of IRE-1 were significantly upregulated in Sestrin2 siRNA group (P < 0.05). Conclusions G-Rg1 protects rat cardiomyocytes and maintains endoplasmic reticulum homeostasis in vitro by upregulating expression of Sestrin2. -
Key words:
- Ginsenoside-rg1 /
- ATF-6 /
- PERK /
- IRE-1 /
- Sestrin2
-

图 1 各组心肌细胞 ROS 水平
用 ROS 阳性细胞所占百分比表示各组心肌细胞中 ROS 水平。**P < 0.01。
Figure 1. The levels of ROS in cardiomyocytes

图 2 各组心肌细胞 HIF-1α、Sestrin2 蛋白表达及内质网跨膜蛋白:ATF-6、PERK、IRE-1 表达
*P < 0.05,**P < 0.01。
Figure 2. The protein expression of HIF-1α,Sestrin2,ATF-6,PERK,IRE-1 in cardiomyocytes
-
[1] Lu M L,Wang J,Sun Y,et al. Ginsenoside Rg1 attenuates mechanical stress-induced cardiac injury via calcium sensing receptor-related pathway[J]. J Ginseng Res,2021,45(6):683-694. doi: 10.1016/j.jgr.2021.03.006 [2] Yang C,Jiang G,Xing Y. Protective effect of ginsenosides Rg1 on ischemic injury of cardiomyocytes after acute myocardial infarction[J]. Cardiovasc Toxicol,2022,22(10-11):910-915. doi: 10.1007/s12012-022-09767-1 [3] Che X,Chai J,Fang Y,et al. Sestrin2 in hypoxia and hypoxia-related diseases[J]. Redox Rep,2021,26(1):111-116. doi: 10.1080/13510002.2021.1948774 [4] Zhu H,Zhou H. Novel insight into the role of endoplasmic reticulum stress in the pathogenesis of myocardial ischemia-reperfusion injury[J]. Oxid Med Cell Longev,2021,2021:5529810. [5] Gao A,Li F,Zhou Q,Chen L. Sestrin2 as a potential therapeutic target for cardiovascular diseases[J]. Pharmacol Res,2020,159:104990. doi: 10.1016/j.phrs.2020.104990 [6] Kishimoto Y,Aoyama M,Saita E,et al. Association between plasma sestrin2 levels and the presence and severity of coronary artery disease[J]. Dis Markers,2020,2020:7439574. [7] Fang C,Yang Z,Shi L,et al. Circulating sestrin levels are increased in hypertension patients[J]. Dis Markers,2020,2020:3787295. [8] Kishimoto Y,Saita E,Ohmori R,et al. Plasma sestrin2 concentrations and carotid atherosclerosis[J]. Clin Chim Acta,2020,504:56-59. doi: 10.1016/j.cca.2020.01.020 [9] Li L,Pan C S,Yan L,et al. Ginsenoside Rg1 ameliorates rat myocardial ischemia-reperfusion injury by modulating energy metabolism pathways[J]. Front Physiol,2018,9:78. doi: 10.3389/fphys.2018.00078 [10] Lyu T J,Zhang Z X,Chen J,Liu Z J. Ginsenoside Rg1 ameliorates apoptosis,senescence and oxidative stress in ox-LDL-induced vascular endothelial cells via the AMPK/SIRT3/p53 signaling pathway[J]. Exp Ther Med,2022,24(3):545. doi: 10.3892/etm.2022.11482 [11] Granger D N,Kvietys P R. Reperfusion injury and reactive oxygen species: The evolution of a concept[J]. Redox Biol,2015,6:524-551. doi: 10.1016/j.redox.2015.08.020 [12] Martindale J J,Fernandez R,Thuerauf D,et al. Endoplasmic reticulum stress gene induction and protection from ischemia/reperfusion injury in the hearts of transgenic mice with a tamoxifen-regulated form of ATF6[J]. Circ Res,2006,98(9):1186-1193. doi: 10.1161/01.RES.0000220643.65941.8d [13] Ruan Y,Zeng J,Jin Q,et al. Endoplasmic reticulum stress serves an important role in cardiac ischemia/reperfusion injury (Review)[J]. Exp Ther Med,2020,20(6):268. [14] Liu Y,Li M,Sun M,et al. Sestrin2 is an endogenous antioxidant that improves contractile function in the heart during exposure to ischemia and reperfusion stress[J]. Free Radic Biol Med,2021,165:385-394. doi: 10.1016/j.freeradbiomed.2021.01.048 [15] Park H W,Park H,Ro S H,et al. Hepatoprotective role of Sestrin2 against chronic ER stress[J]. Nat Commun,2014,5:4233. doi: 10.1038/ncomms5233 [16] Wang L X,Zhu X M,Luo Y N,et al. Sestrin2 protects dendritic cells against endoplasmic reticulum stress-related apoptosis induced by high mobility group box-1 protein[J]. Cell Death Dis,2020,11(2):125. doi: 10.1038/s41419-020-2324-4 [17] Zheng J,Chen P,Zhong J,et al. HIF-1α in myocardial ischemia-reperfusion injury (Review)[J]. Mol Med Rep,2021,23(5):352. [18] Essler S,Dehne N,Brüne B. Role of sestrin2 in peroxide signaling in macrophages[J]. FEBS Lett,2009,583(21):3531-3535. doi: 10.1016/j.febslet.2009.10.017 [19] Ala M. Sestrin2 in cancer: a foe or a friend[J]. Biomark Res,2022,10(1):29. doi: 10.1186/s40364-022-00380-6 [20] Lu C,Jiang Y,Xu W,Bao X. Sestrin2: multifaceted functions,molecular basis,and its implications in liver diseases[J]. Cell Death Dis,2023,14(2):160. doi: 10.1038/s41419-023-05669-4 -

 下载:
下载:
 点击查看大图
点击查看大图
图(3)
计量
- 文章访问数: 3293
- HTML全文浏览量: 1922
- PDF下载量: 9
- 被引次数: 0
