

Effect of Bifidobacterium trisporus on Intestinal Microbiota in the Mouse Model of Non-alcoholic Fatty Liver Disease
-
摘要:
目的 探究双歧杆菌三联活菌在小鼠NAFLD模型中的疗效和对肠道微生物组的影响。 方法 使用C57BL/6J小鼠分为3个实验组别,对照组(n = 10):小鼠使用普通饮食饲养;NAFLD模型组(n = 10):使用高脂饮食饲养;双歧杆菌组(n = 10):使用NAFLD模型小鼠并给予双歧杆菌三联活菌治疗。在实验结束时间点对不同组小鼠肝脏石蜡切片进行HE染色观察病理变化。采用16S rRNA基因测序技术完成对各组小鼠肠道菌群的检测,并进行生物信息学分析。 结果 与对照组相比,NAFLD小鼠肝脏脂肪累积增加,脂滴增多,细胞肥大,并且血清谷丙转氨酶(ALT)水平增加(P < 0.0001);经双歧杆菌治疗后,NAFLD小鼠肝脏脂肪积累减少并且血清ALT水平降低(P < 0.0001)。肠道微生物群分析在3个组别中鉴定出共有菌种348种。微生物功能分析显示,双歧杆菌组肠道微生物代谢功能水平介于对照组和NAFLD组之间;双歧杆菌治疗能改变NAFLD模型中特定肠道微生物的丰度变化,并且这些肠道微生物可能参与了谷胱甘肽代谢、嘌呤代谢和鞘脂代谢等途径调节机体的代谢功能。 结论 双歧杆菌三联活菌在小鼠模型中可能通过调节肠道微生物组的功能改善NAFLD,并且双歧杆菌三联活菌有改善机体脂代谢紊乱的潜力。 -
关键词:
- 非酒精性脂肪性肝(NAFLD) /
- 宏基因组 /
- 双歧杆菌三联活菌 /
- 高脂饮食
Abstract:Objective To investigate the therapeutic effects of probiotics containing Bifidobacterium on a mouse model of non-alcoholic fatty liver disease (NAFLD) and its impact on the gut microbiota. Methods C57BL/6J mice were divided into three experimental groups. The control group (n = 10) of mice was fed a normal diet, the NAFLD model group (n = 10) was fed a high-fat diet, and the probiotics group (n = 10) was given Bifidobacteriumtrisporus treatment while using NAFLD model mice. HE staining was performed on paraffin sections of mouse liver from different groups at the end of the experiment to observe pathological changes. The 16S rRNA gene sequencing technique was used to detect the gut microbiota in each group of mice, followed by bioinformatics analysis. Results Compared with the control group, NAFLD mice showed increased accumulation of liver fat, increased number of lipid droplets, enlarged cells, and increased levels of serum alanine aminotransferase (ALT)(P < 0.0001). After treatment with Bifidobacteriumtrisporus, the accumulation of liver fat in NAFLD mice decreased and serum ALT levels decreased (P < 0.0001). Microbiota analysis of the intestinal tract identified 348 common species of microorganisms in the three groups. Functional analysis of the microbiota showed that the metabolic function of the intestinal microbiota in the Bifidobacterium group was intermediate between the control group and the NAFLD group; Bifidobacterium treatment was able to alter the abundance of specific intestinal microorganisms in the NAFLD model, and these intestinal microorganisms may be involved in the regulation of metabolic function pathways such as glutathione metabolism, purine metabolism, and sphingolipid metabolism in the body. Conclusion Bifidobacterium may improve NAFLD by regulating the function of the intestinal microbiota in a mouse model, and it has the potential to improve disordered lipid metabolism in the body. -

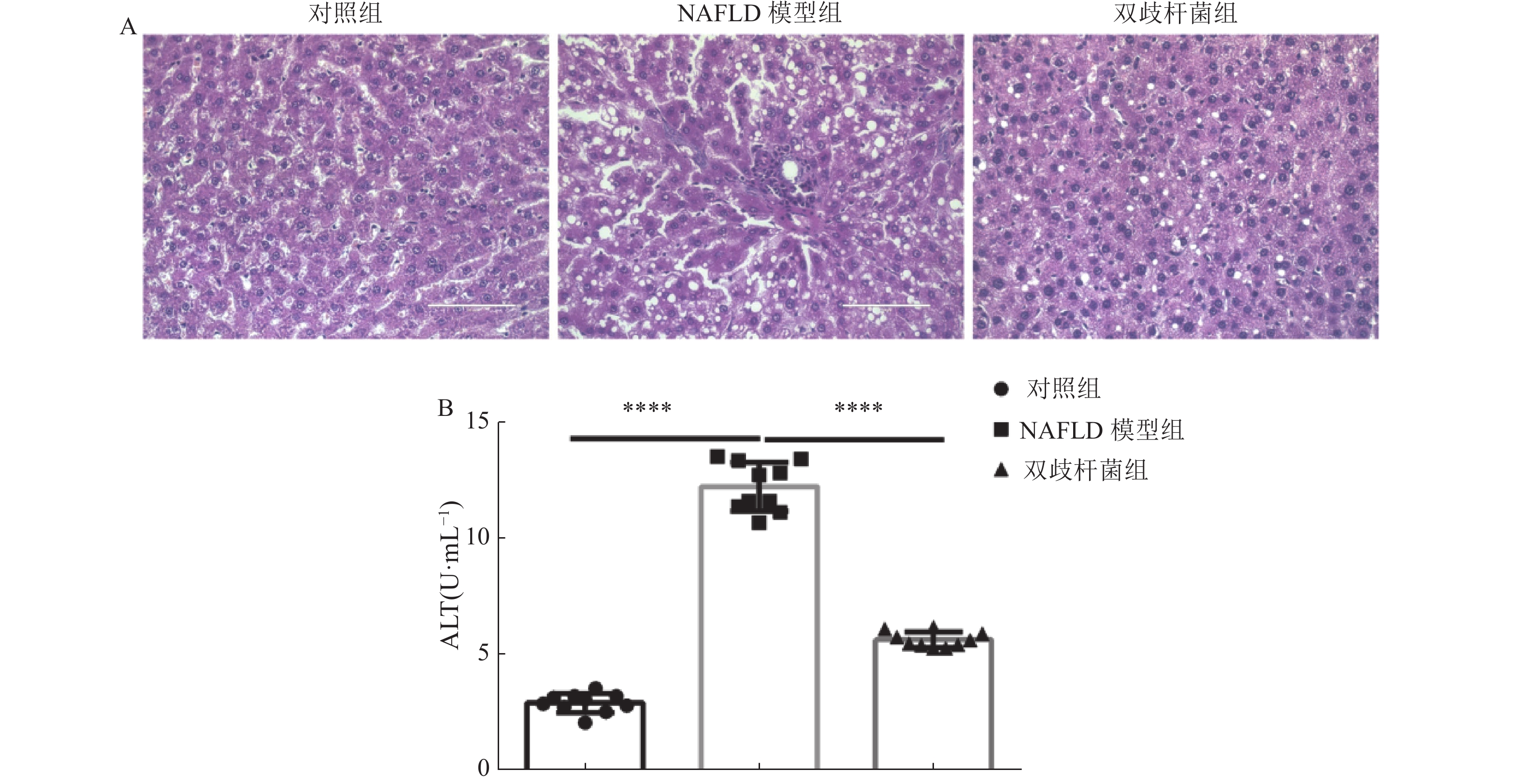
图 1 双歧杆菌改善小鼠NAFLD发生
A:对照组,NAFLD组和双歧杆菌组小鼠肝脏HE染色(200×);B:对照组, NAFLD组和双歧杆菌组小鼠血清中谷丙转氨酶(ALT)活性检测。对照组与 NAFLD组ALT比较,****P < 0.0001;NAFLD组和双歧杆菌组ALT比较,****P < 0.0001。
Figure 1. Bifidobacterium improves the NAFLD in the mouse model

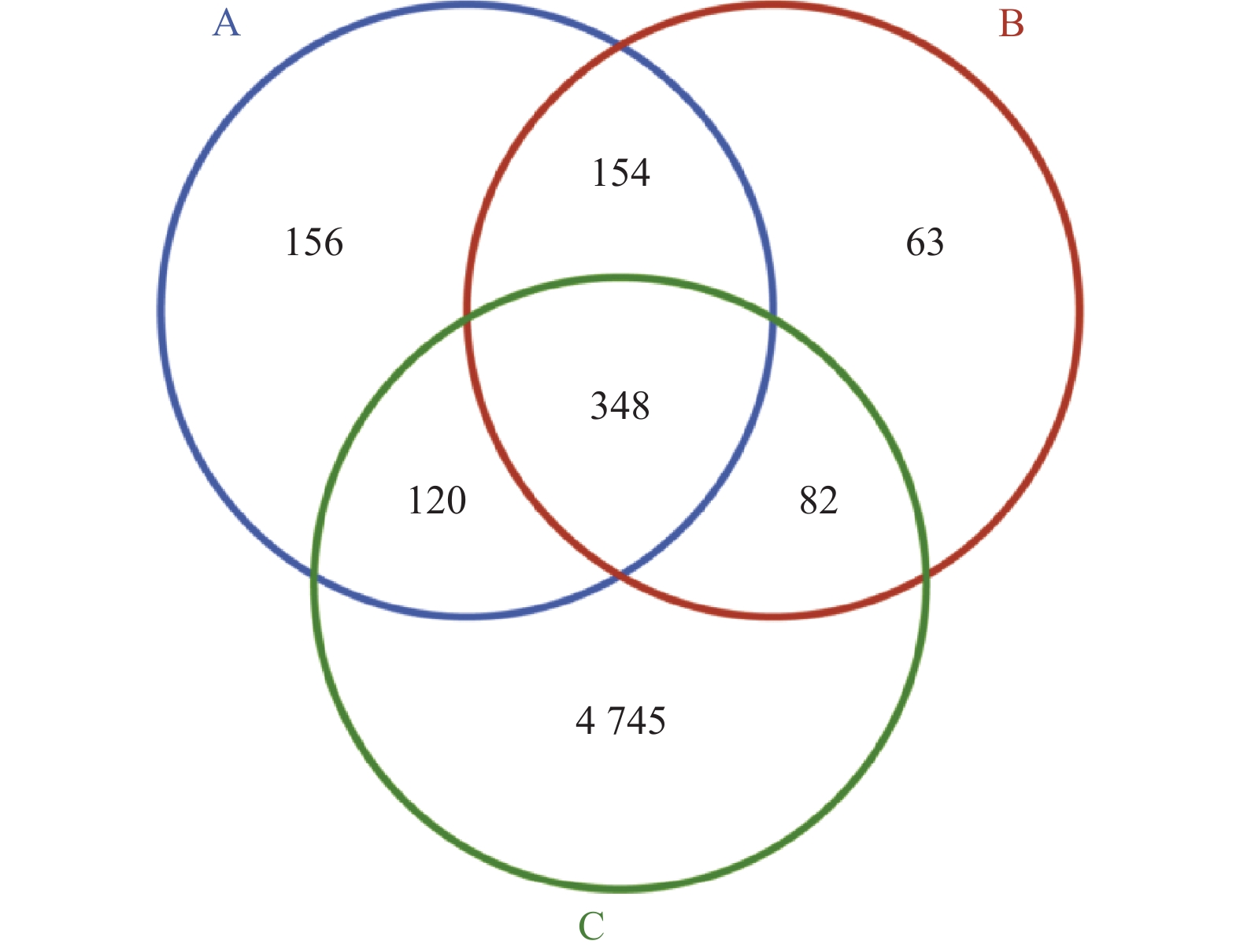
图 2 各组小鼠粪便样本鉴定微生物种类韦恩图
A:对照组;B:NAFLD组;C:双歧杆菌组。
Figure 2. Venn diagram of microbial species identified from fecal samples of mice in each group

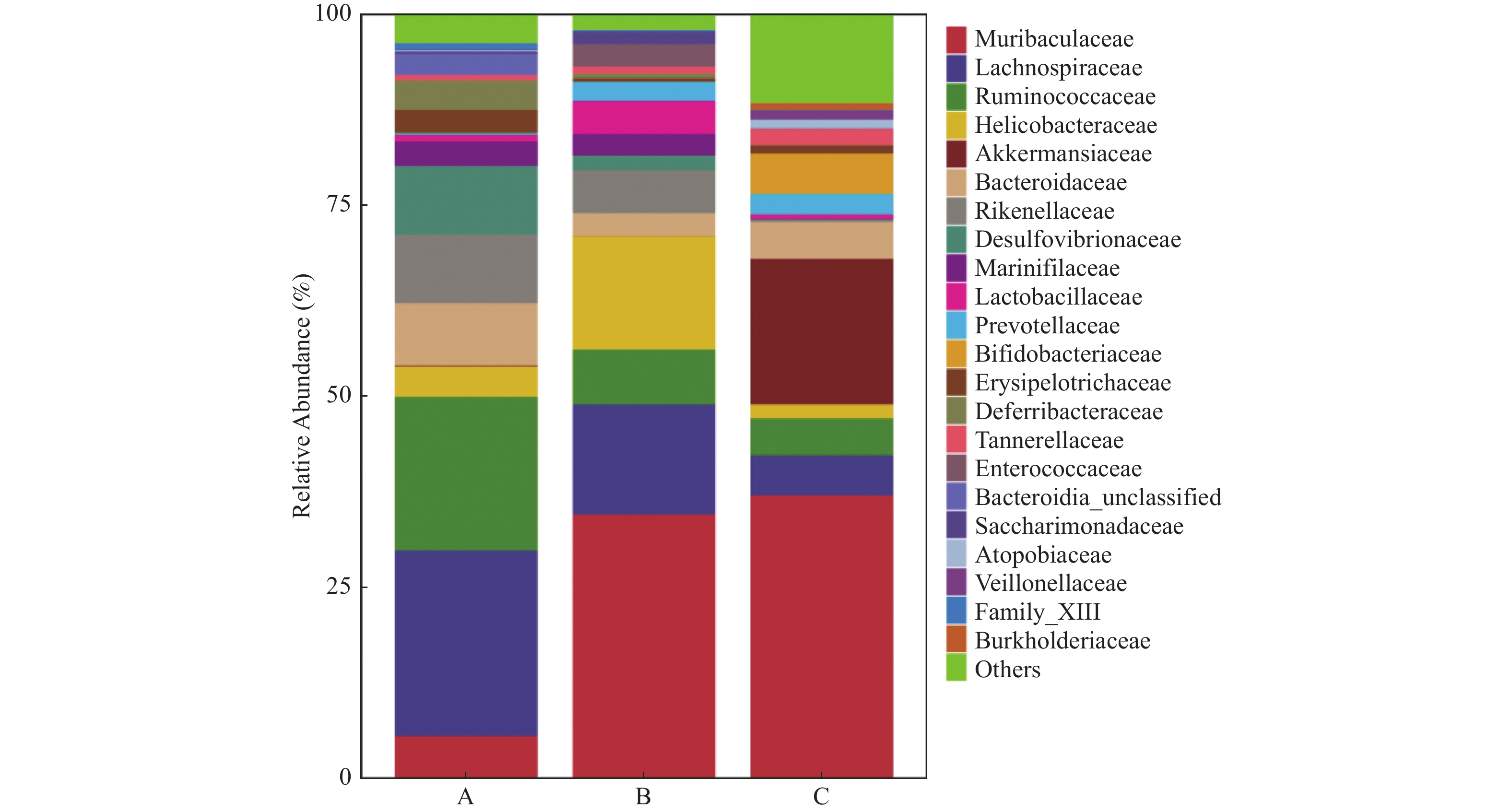
图 3 各组小鼠粪便样本鉴定微生物种类丰度分布
A:对照组;B:NAFLD组;C:双歧杆菌组。
Figure 3. Abundance distribution of identified microbial species in fecal samples in each group

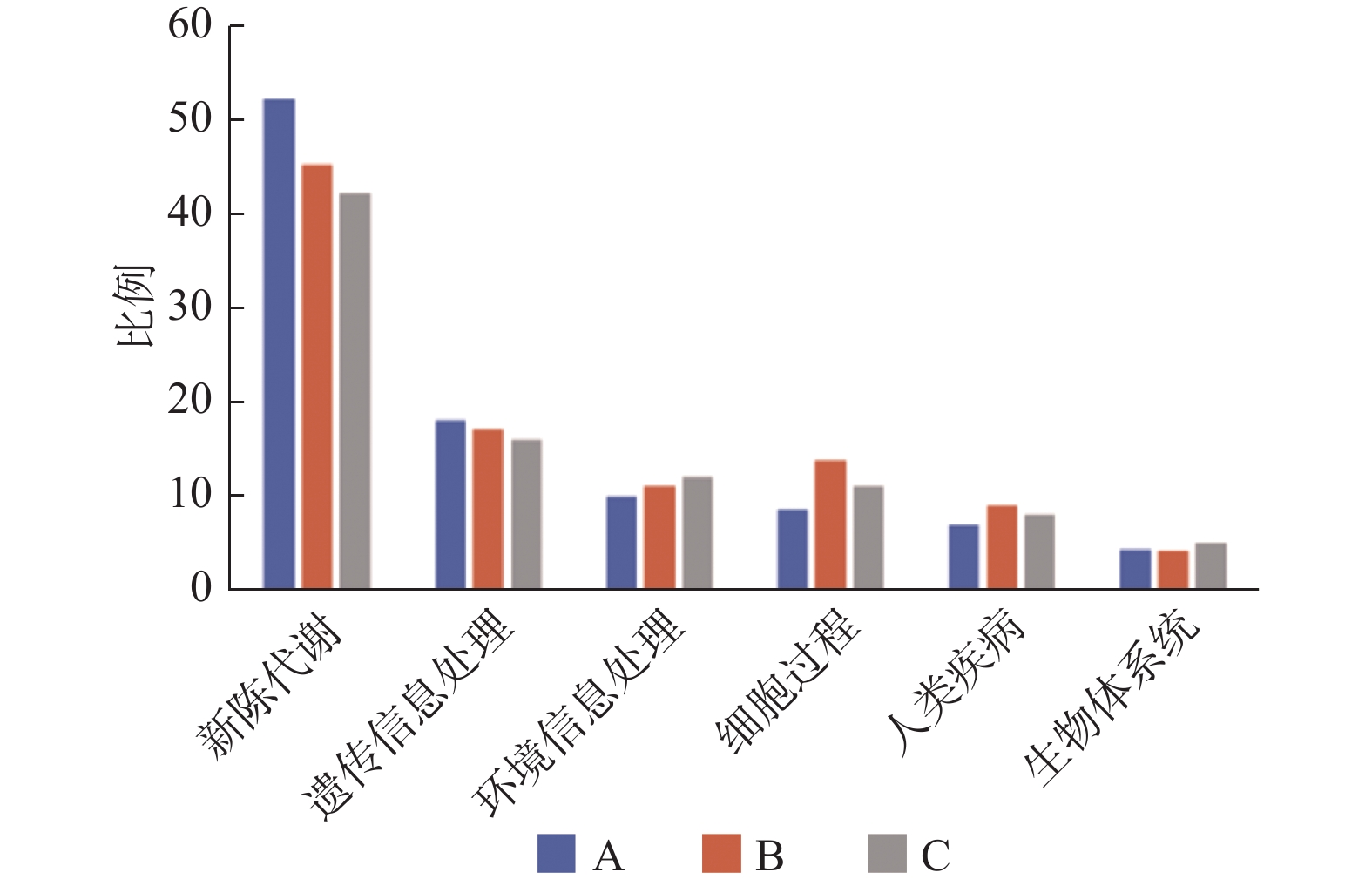
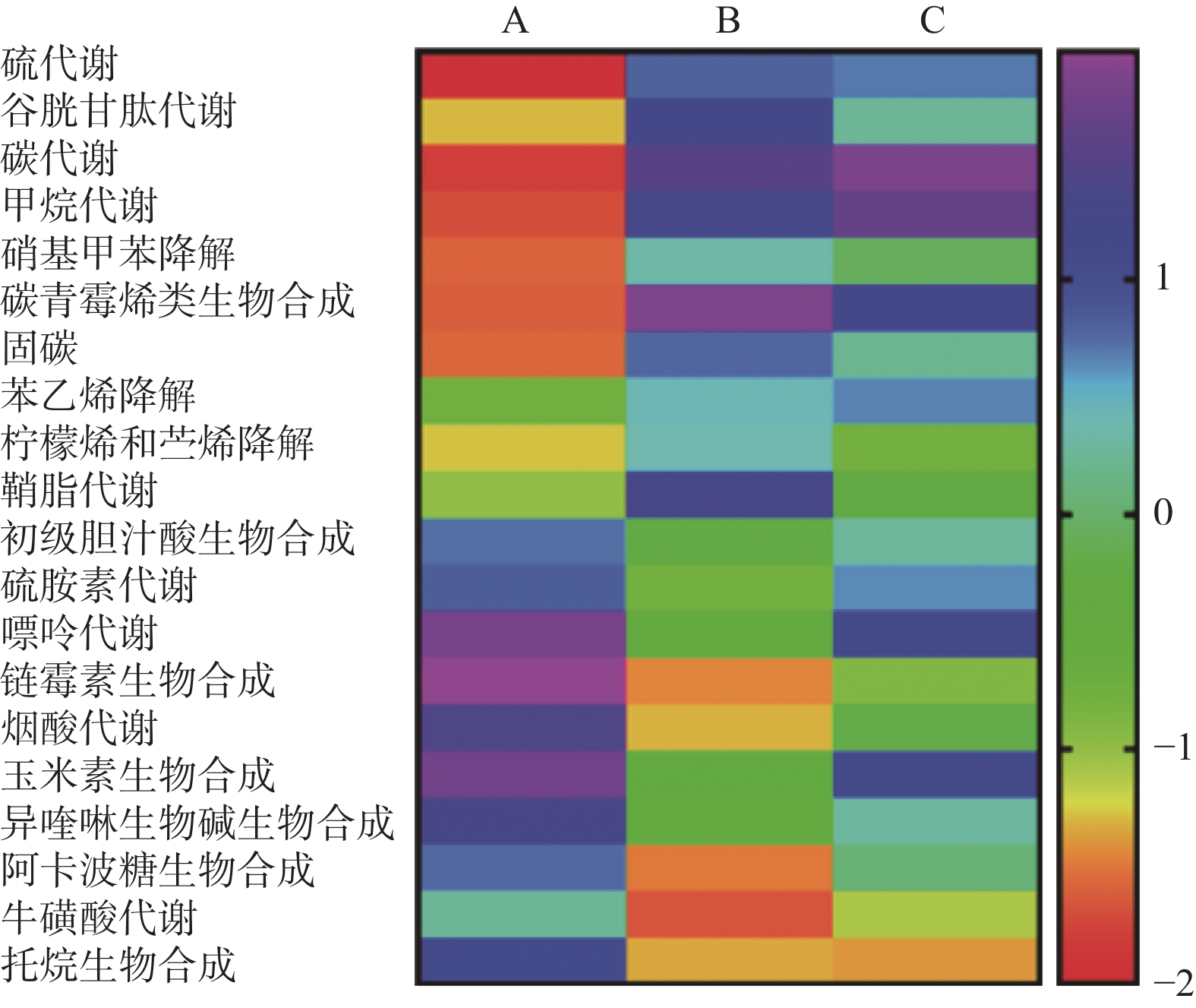
图 4 各组小鼠粪便样本宏基因组数据测序基因片段功能富集分析
A:对照组;B:NAFLD组;C:双歧杆菌组。
Figure 4. Functional enrichment analysis of gene fragments in the metagenomic data of fecal samples from mice in each group
-
[1] Wagner R,Eckstein S S,Yamazaki H,et al. Metabolic implications of pancreatic fat accumulation[J]. Nat Rev Endocrinol,2022,18(1):43-54. doi: 10.1038/s41574-021-00573-3 [2] Kwan S Y,Jiao J,Joon A,et al. Gut microbiome features associated with liver fibrosis in Hispanics,a population at high risk for fatty liver disease[J]. Hepatology,2022,5(4):955-967. [3] Kolodziejczyk A A,Zheng D,Shibolet O,et al. The role of the microbiome in NAFLD and NASH[J]. EMBO Mol Med,2019,11(2):e9302. doi: 10.15252/emmm.201809302 [4] Safari Z,Gérard P. The links between the gut microbiome and non-alcoholic fatty liver disease (NAFLD)[J]. Cell Mol Life Sci,2019,76(8):1541-1558. doi: 10.1007/s00018-019-03011-w [5] Hu H,Lin A,Kong M,et al. Intestinal microbiome and NAFLD: Molecular insights and therapeutic perspectives[J]. J Gastroenterol,2020,55(2):142-158. doi: 10.1007/s00535-019-01649-8 [6] Aron-Wisnewsky J,Vigliotti C,Witjes J,et al. Gut microbiota and human NAFLD: Disentangling microbial signatures from metabolic disorders[J]. Nat Rev Gastroenterol Hepatol,2020,17(5):279-297. doi: 10.1038/s41575-020-0269-9 [7] Lau L H S,Wong S H. Microbiota,obesity and NAFLD[J]. Adv Exp Med Biol,2018,1061:111-125. [8] Canfora E E,Meex R C R,Venema K,et al. Gut microbial metabolites in obesity,NAFLD and T2DM[J]. Nat Rev Endocrinol,2019,15(5):261-273. doi: 10.1038/s41574-019-0156-z [9] Fan Y,Pedersen O. Gut microbiota in human metabolic health and disease[J]. Nat Rev Microbiol,2021,19(1):55-71. doi: 10.1038/s41579-020-0433-9 [10] Bana B,Cabreiro F. The microbiome and aging[J]. Annu Rev Genet,2019,53:239-261. doi: 10.1146/annurev-genet-112618-043650 [11] Suez J,Zmora N,Segal E,et al. The pros,cons,and many unknowns of probiotics[J]. Nat Med,2019,25(5):716-729. doi: 10.1038/s41591-019-0439-x [12] Qian X,Si Q,Lin G,et al. Bifidobacterium adolescentis is effective in relieving type 2 diabetes and may be related to its dominant core genome and gut microbiota modulation capacity[J]. Nutrients,2022,14(12):2479. doi: 10.3390/nu14122479 [13] Fang Z,Pan T,Li L,et al. Bifidobacterium longum mediated tryptophan metabolism to improve atopic dermatitis via the gut-skin axis[J]. Gut Microbes,2022,14(1):2044723. doi: 10.1080/19490976.2022.2044723 [14] Eng J M,Estall J L. Diet-induced models of non-alcoholic fatty liver disease: Food for thought on sugar,fat,and cholesterol[J]. Cells,2021,10(7):1805.P. doi: 10.3390/cells10071805 [15] Zeng H,Larson K J,Cheng WH,et al. Advanced liver steatosis accompanies an increase in hepatic inflammation,colonic,secondary bile acids and Lactobacillaceae/Lachnospiraceae bacteria in C57BL/6 mice fed a high-fat diet[J]. J Nutr Biochem,2020,78:108336. doi: 10.1016/j.jnutbio.2019.108336 [16] Yuan G,Tan M,Chen X. Punicic acid ameliorates obesity and liver steatosis by regulating gut microbiota composition in mice[J]. Food Funct,2021,12(17):7897-7908. doi: 10.1039/D1FO01152A [17] Wang H,Wang Q,Yang C,et al. Bacteroides acidifaciens in the gut plays a protective role against CD95-mediated liver injury[J]. Gut Microbes,2022,14(1):2027853. doi: 10.1080/19490976.2022.2027853 [18] Nishimura N,Kaji K,Kitagawa K,et al. Intestinal permeability is a mechanical rheostat in the pathogenesis of liver cirrhosis[J]. Int J Mol Sci,2021,22(13):6921. doi: 10.3390/ijms22136921 [19] Do M H,Oh M J,Lee H B,et al. Bifidobacterium animalis ssp. lactis MG741 reduces body weight and ameliorates nonalcoholic fatty liver disease via improving the gut permeability and amelioration of inflammatory cytokines[J]. Nutrients,2022,14(9):1965. doi: 10.3390/nu14091965 [20] Wang W,Xu A L,Li Z C,et al. Combination of probiotics and salvia miltiorrhiza polysaccharide alleviates hepatic steatosis via gut microbiota modulation and insulin resistance improvement in high fat-induced NAFLD mice[J]. Diabetes Metab J,2020,44(2):336-348. doi: 10.4093/dmj.2019.0042 [21] Yang X,Mo W,Zheng C,et al. Alleviating effects of noni fruit polysaccharide on hepatic oxidative stress and inflammation in rats under a high-fat diet and its possible mechanisms[J]. Food Funct,2020,11(4):2953-2968. doi: 10.1039/D0FO00178C [22] Wang X,Shi L,Wang X,et al. MDG-1,an Ophiopogon polysaccharide,restrains process of non-alcoholic fatty liver disease via modulating the gut-liver axis[J]. Int J Biol Macromol,2019,141(3):1013-1021. [23] Tong A J,Hu R K,Wu L X,et al. Ganoderma polysaccharide and chitosan synergistically ameliorate lipid metabolic disorders and modulate gut microbiota composition in high fat diet-fed golden hamsters[J]. J Food Biochem,2020,44(1):e13109. [24] Dehhaghi M, Kazemi Shariat Panahi H, Guillemin GJ. Microorganisms, tryptophan metabolism, and kynurenine pathway: A complex interconnected loop influencing human health status [J]. Int J Tryptophan Res, 2019, 12(2): 1178646919852996. [25] Arnoriaga-Rodríguez M,Mayneris-Perxachs J,Burokas A,et al. Obesity impairs short-term and working memory through gut microbial metabolism of aromatic amino acids[J]. Cell Metab,2020,32(4):548-560,e7. doi: 10.1016/j.cmet.2020.09.002 [26] Zhao Z H,Xin F Z,Xue Y,et al. Indole-3-propionic acid inhibits gut dysbiosis and endotoxin leakage to attenuate steatohepatitis in rats[J]. Exp Mol Med,2019,51(9):1-14. -

 下载:
下载:
 点击查看大图
点击查看大图
计量
- 文章访问数: 2013
- HTML全文浏览量: 1199
- PDF下载量: 32
- 被引次数: 0
