

Artemisinin Mediates the Aerobic Glycolysis of Clear Cell Renal Cell Carcinomas via Regulating ENO2
-
摘要:
目的 探讨青蒿素(Artemisinin)调控ENO2对肾透明细胞癌(clear cell renal cell carcinomas,ccRCC)细胞增殖和有氧糖酵解的作用及具体调控机制。 方法 分别将ccRCC细胞系OSRC2和ACHN暴露于0、10、20、30、40 μmol/L青蒿素中,OSRC2和ACHN细胞经25 μmol/L青蒿素处理或同时转染si-ENO2,CCK-8检测细胞活力,葡萄糖测试盒和乳酸测试盒检测葡萄糖消耗和乳酸生成量,Western blot检测HK2、LDHA和ENO2的表达,RT-qPCR检测ENO2 mRNA相对表达。 结果 OSRC2和ACHN细胞存活率随青蒿素浓度和暴露时间的增加而降低,OSRC2细胞的IC50值为25.47 μmol/L,ACHN细胞的IC50值为26.31 μmol/L。ENO2在OSRC2和ACHN细胞中表达升高(P < 0.01),青蒿素可下调ENO2在癌细胞中的表达(P < 0.05)。暴露于25 μmol/L青蒿素或敲降ENO2可抑制OSRC2和ACHN细胞的存活(P < 0.001)、葡萄糖消耗(P < 0.05)、乳酸生成(均P < 0.05)以及HK2和LDHA的蛋白表达(P < 0.05)。同时敲降ENO2且暴露于25 μmol/L青蒿素组中OSRC2和ACHN细胞的存活率(P < 0.001)、葡萄糖消耗(P < 0.05)、乳酸生成(P < 0.05)以及HK2和LDHA的表达(均P < 0.05)低于仅敲降ENO2组。 结论 青蒿素可抑制ccRCC细胞的存活率和有氧糖酵解,并通过下调ENO2在ccRCC中的表达而发挥作用。 Abstract:Objective To investigate Artemisinin-affected cell proliferation and aerobic glycolysis on clear cell renal carcinomas(ccRCC) through regulating ENO2 and clarify the mechanism. Methods ccRCC cell lines OSRC2 and ACHN were exposed to 0, 10, 20, 30, and 40 μmol/L Artemisinin, OSRC2 and ACHN cells treated with 25 μmol/L Artemisinin or transfected with si-ENO2 at the same time. CCK-8 assay detected cell proliferation, glucose consumption, and lactate production were detected by glucose test kit and lactate test kit. The expressions of HK2, LDHA, and ENO2 were detected by Western blot, and ENO2 mRNA expression was detected by RT-qPCR. Results The survival rate of OSRC2 and ACHN cells were decreased with the increase of Artemisinin concentration and treatment time, the IC50 of OSRC2 cells was 25.47 μmol/L, and the IC50 of ACHN was 26.31 μmol/L. ENO2 was upregulated in OSRC2 and ACHN cells(P < 0.01), and Artemisinin diminished ENO2 expression in cancer cells(all P < 0.05). Exposure to Artemisinin(25 μmol/L) or knocking down ENO2 significantly inhibited the survival of OSRC2 and ACHN cells(all P < 0.001), glucose consumption(P < 0.05), lactate production(all P < 0.05) and protein expression of HK2 and LDHA(all P < 0.05). Compared with si-ENO2 group, the survival rate(P < 0.001), glucose consumption(P < 0.05), lactate production(all P < 0.05) and the expression of HK2 and LDHA(all P < 0.05) of OSRC2 and ACHN cells in knocking down ENO2 and treated with 25 μmol/L Artemisinin group were decreased. Conclusion Artemisinin can inhibit the survival rate and aerobic glycolysis of ccRCC cells, and plays a role by downregulating the expression of ENO2 in ccRCC. -
Key words:
- Clear cells renal cell carcinomas /
- Aerobic glycolysis /
- Artemisinin /
- ENO2 /
- Survival rate
-

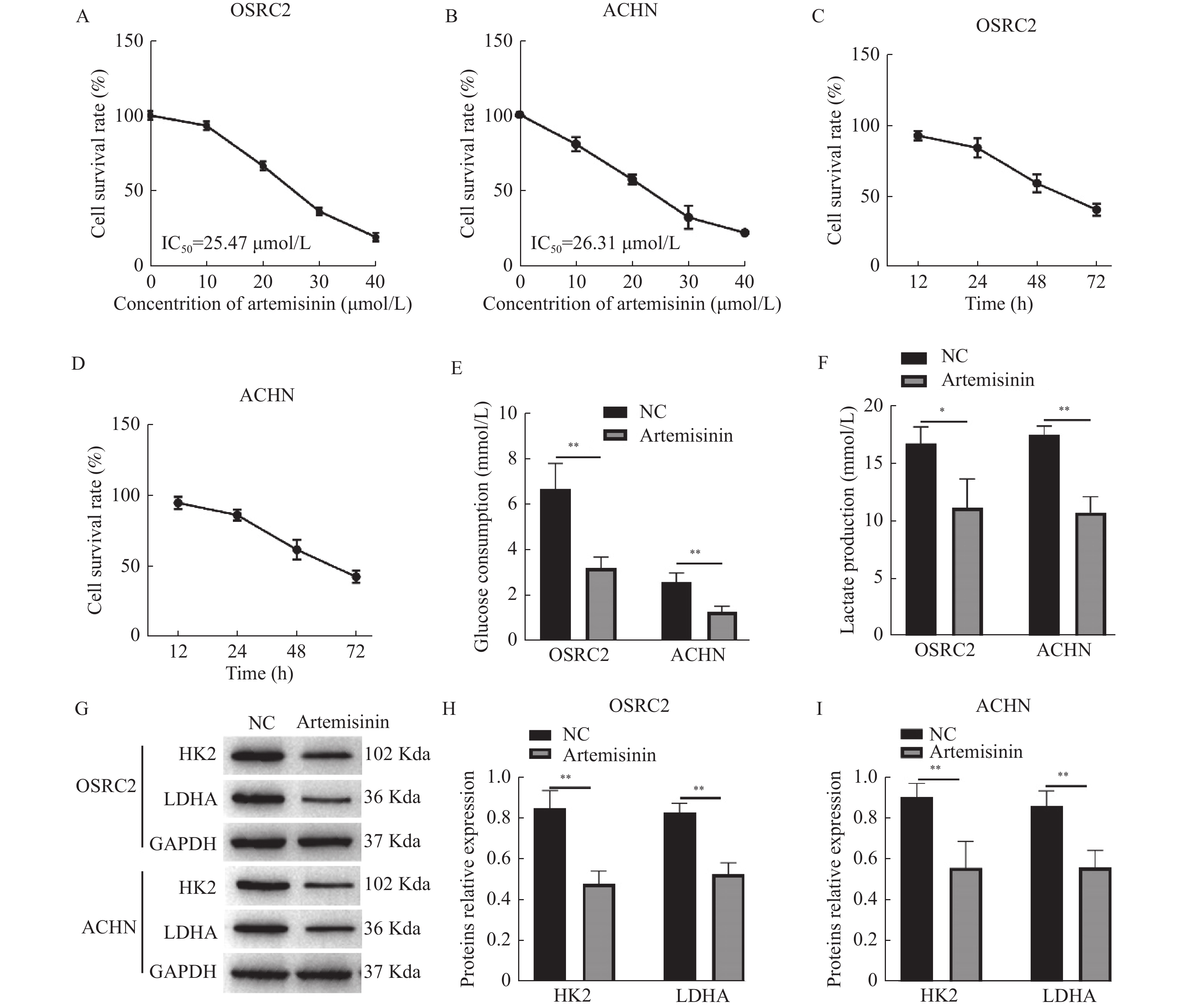
图 1 青蒿素抑制ccRCC细胞存活率和糖酵解
A~B:CCK-8检测不同浓度青蒿素处理时OSRC2和ACHN细胞的存活率;C~D:CCK-8检测25 μmol/L青蒿素处理不同时间对OSRC2和ACHN细胞存活率的作用;E:试剂盒检测葡萄糖消耗量;F:试剂盒检测乳酸生成;G~I:Western blot检测HK2和LDHA的生成。*P < 0.05,**P < 0.01。
Figure 1. Artemisinin repressed the survival rate and glycolysis of ccRCC cells

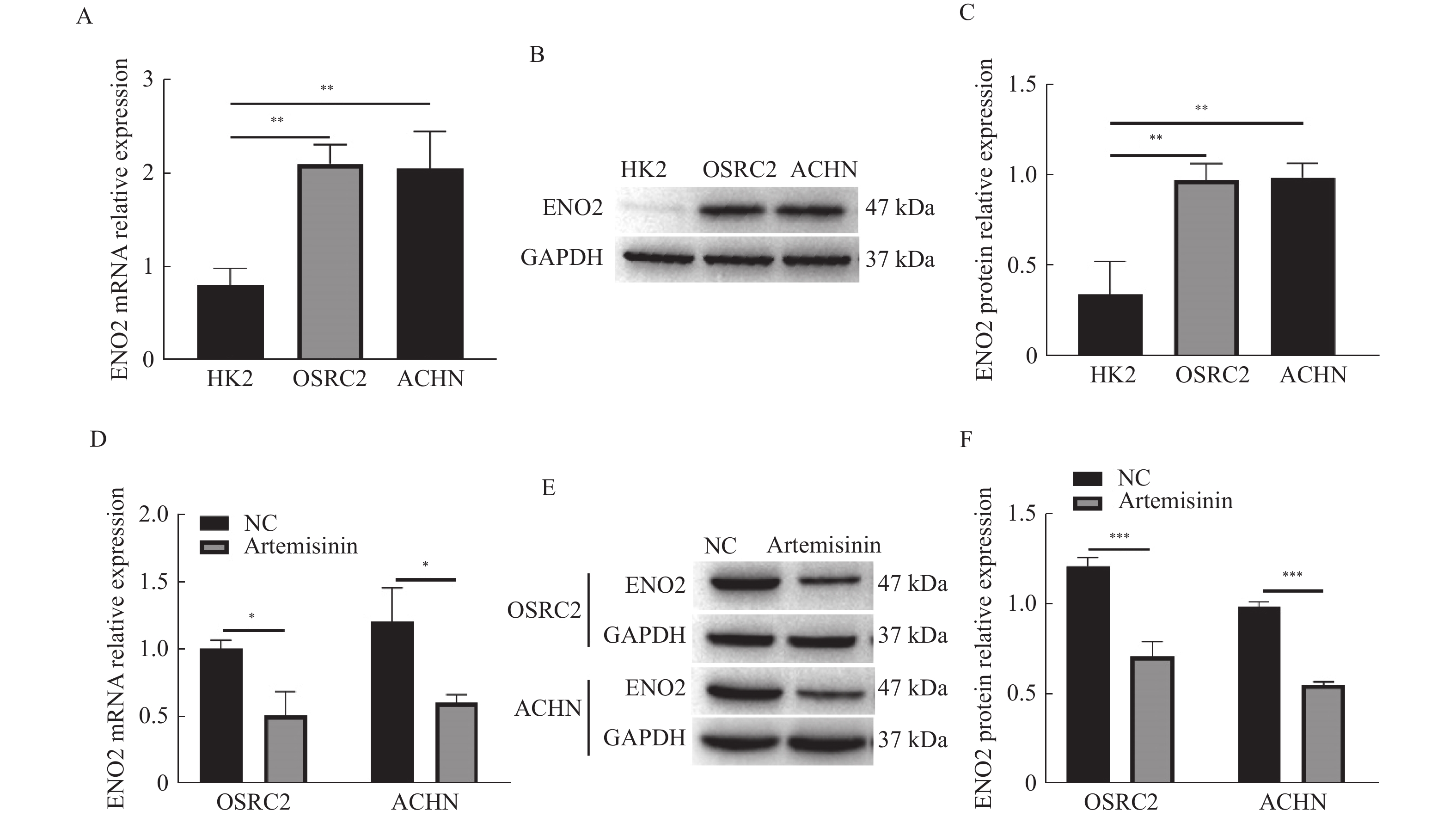
图 2 青蒿素下调ENO2在癌细胞中的表达
A:RT-qPCR检测ENO2 mRNA相对表达;B~C:Western blot检测ENO2蛋白相对表达;D:RT-qPCR检测青蒿素对ENO2 mRNA相对表达的作用;E~F:Western blot检测青蒿素对ENO2蛋白相对表达的作用。*P < 0.05,**P < 0.01,***P < 0.001。
Figure 2. Artemisinin inhibited the expression of ENO2 in ccRCC cells

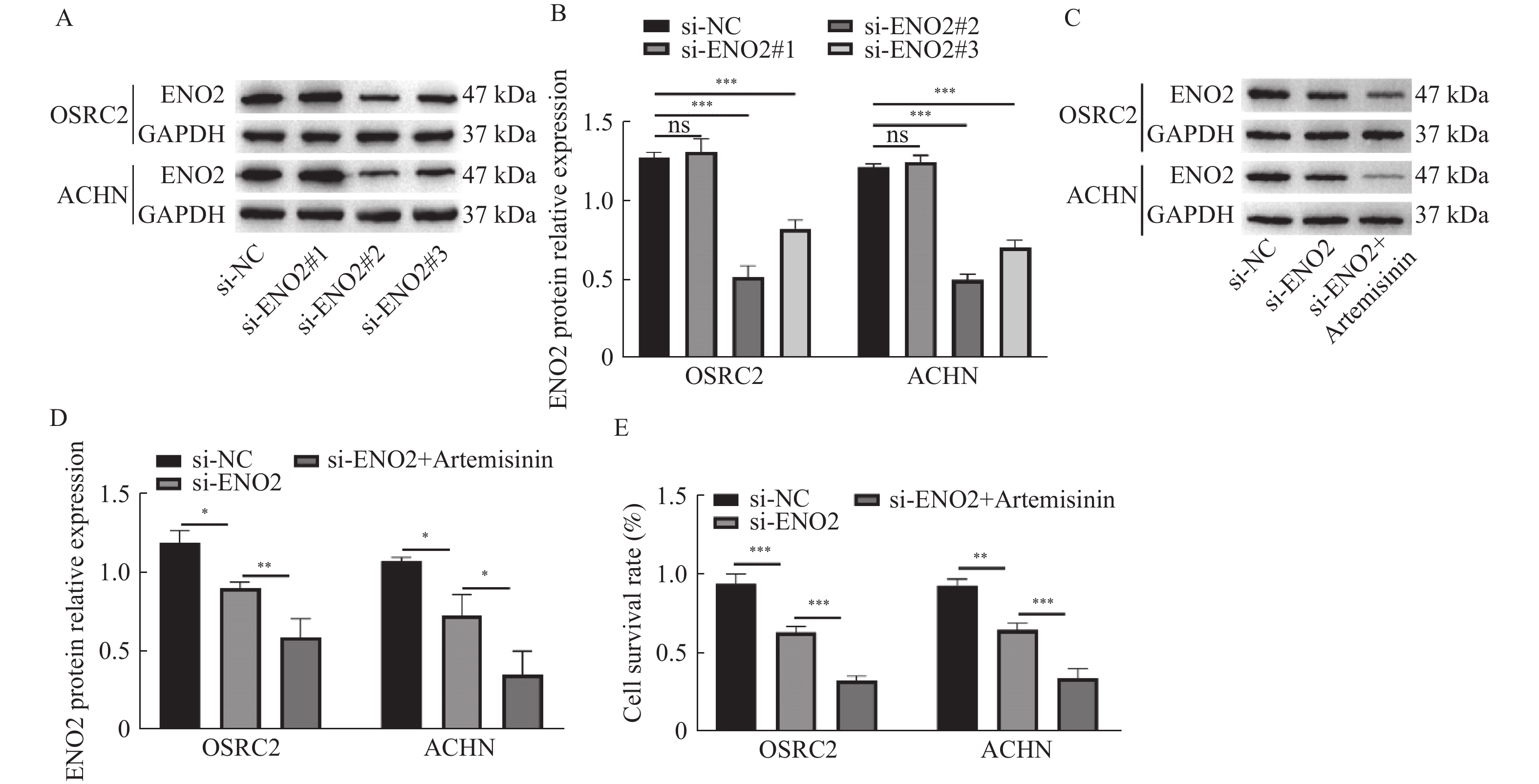
图 3 青蒿素通过下调ENO2抑制癌细胞活力
A~B:Western blot检测si-ENO2转染效率;C~D:Western blot检测ENO2蛋白相对表达;E:CCK-8检测细胞存活率。*P < 0.05,**P < 0.01,***P < 0.001。
Figure 3. Artemisinin repressed cancer cell viability by downregulating ENO2 expression
-
[1] Znaor A,Lortet-Tieulent J,Laversanne M,et al. International variations and trends in renal cell carcinoma incidence and mortality[J]. Eur Urol,2015,67(3):519-530. doi: 10.1016/j.eururo.2014.10.002 [2] Baldewijns M M,Van Vlodrop I J,Schouten L J,et al. Genetics and epigenetics of renal cell cancer[J]. Biochim Biophys Acta,2008,1785(2):133-155. [3] Vaupel P,Schmidberger H,Mayer A. The Warburg effect: Essential part of metabolic reprogramming and central contributor to cancer progression[J]. Int J Radiat Biol,2019,95(7):912-919. doi: 10.1080/09553002.2019.1589653 [4] Nocera G,Jacob C. Mechanisms of Schwann cell plasticity involved in peripheral nerve repair after injury[J]. Cell Mol Life Sci,2020,77(20):3977-3989. doi: 10.1007/s00018-020-03516-9 [5] Reed G H,Poyner R R,Larsen T M,et al. Structural and mechanistic studies of enolase[J]. Curr Opin Struct Biol,1996,6(6):736-743. doi: 10.1016/S0959-440X(96)80002-9 [6] Chen W J,Yang W,Gong M,et al. ENO2 affects the EMT process of renal cell carcinoma and participates in the regulation of the immune microenvironment[J]. Oncol Rep,2023,49(2):33. [7] Tang C,Wang M,Dai Y,et al. Krüppel-like factor 12 suppresses bladder cancer growth through transcriptionally inhibition of enolase 2[J]. Gene,2021,769:145338. [8] Fang L,Ye T,An Y. Circular RNA FOXP1 induced by ZNF263 upregulates U2AF2 expression to accelerate renal cell carcinoma tumorigenesis and warburg effect through sponging miR-423-5p[J]. J Immunol Res,2021,2021:8050993. [9] Wang M,Chen H,He X,et al. Artemisinin inhibits the development of esophageal cancer by targeting HIF-1α to reduce glycolysis levels[J]. J Gastrointest Oncol,2022,13(5):2144-2153. doi: 10.21037/jgo-22-877 [10] Farhan M,Silva M,Xingan X,et al. Artemisinin inhibits the migration and invasion in uveal melanoma via inhibition of the PI3K/AKT/mTOR signaling pathway[J]. Oxid Med Cell Longev,2021,2021:9911537. [11] Wang Z,Li M,Liu Y,et al. Dihydroartemisinin triggers ferroptosis in primary liver cancer cells by promoting and unfolded protein response-induced upregulation of CHAC1 expression[J]. Oncol Rep,2021,46(5):240. doi: 10.3892/or.2021.8191 [12] Cai X,Miao J,Sun R,et al. Dihydroartemisinin overcomes the resistance to osimertinib in EGFR-mutant non-small-cell lung cancer[J]. Pharmacol Res,2021,170:105701. [13] Yu C,Sun P,Zhou Y,et al. Inhibition of AKT enhances the anti-cancer effects of Artemisinin in clear cell renal cell carcinoma[J]. Biomed Pharmacother,2019,118:109383. [14] Zhang M X,Wang J L,Mo C Q,et al. CircME1 promotes aerobic glycolysis and sunitinib resistance of clear cell renal cell carcinoma through cis-regulation of ME1[J]. Oncogene,2022,41(33):3979-3990. doi: 10.1038/s41388-022-02386-8 [15] He Y,Wang X,Lu W,et al. PGK1 contributes to tumorigenesis and sorafenib resistance of renal clear cell carcinoma via activating CXCR4/ERK signaling pathway and accelerating glycolysis[J]. Cell Death Dis,2022,13(2):118. doi: 10.1038/s41419-022-04576-4 [16] Li J,Zhang S,Liao D,et al. Overexpression of PFKFB3 promotes cell glycolysis and proliferation in renal cell carcinoma[J]. BMC Cancer,2022,22(1):83. doi: 10.1186/s12885-022-09183-2 [17] Chen X,Li Z,Yong H,et al. Trim21-mediated HIF-1α degradation attenuates aerobic glycolysis to inhibit renal cancer tumorigenesis and metastasis[J]. Cancer Lett,2021,508:115-126. doi: 10.1016/j.canlet.2021.03.023 [18] Gao L,Yang F,Tang D,et al. Mediation of PKM2-dependent glycolytic and non-glycolytic pathways by ENO2 in head and neck cancer development[J]. J Exp Clin Cancer Res,2023,42(1):1. doi: 10.1186/s13046-022-02574-0 [19] Liu D,Mao Y,Chen C,et al. Expression patterns and clinical significances of ENO2 in lung cancer: an analysis based on Oncomine database[J]. Ann Transl Med,2020,8(10):639. doi: 10.21037/atm-20-3354 [20] Soh M A,Garrett S H,Somji S,et al. Arsenic,cadmium and neuron specific enolase (ENO2,γ-enolase) expression in breast cancer[J]. Cancer Cell Int,2011,11(1):41. doi: 10.1186/1475-2867-11-41 [21] Huang J,Yang M,Liu Z,et al. PPFIA4 promotes colon cancer cell proliferation and migration by enhancing tumor glycolysis[J]. Front Oncol,2021,11:653200. [22] Peng J,Liu F,Zheng H,et al. IncRNA ZFAS1 contributes to the radioresistance of nasopharyngeal carcinoma cells by sponging hsa-miR-7-5p to upregulate ENO2[J]. Cell Cycle,2021,20(1):126-141. doi: 10.1080/15384101.2020.1864128 [23] Liu C C,Wang H,Wang W D,et al. ENO2 promotes cell proliferation,glycolysis,and glucocorticoid-resistance in acute lymphoblastic leukemia[J]. Cell Physiol Biochem,2018,46(4):1525-1535. doi: 10.1159/000489196 [24] Pan J,Jin Y,Xu X,et al. Integrated analysis of the role of enolase 2 in clear cell renal cell carcinoma[J]. Dis Markers,2022,2022:6539203. -

 下载:
下载:

 点击查看大图
点击查看大图
计量
- 文章访问数: 1970
- HTML全文浏览量: 1077
- PDF下载量: 29
- 被引次数: 0
