

The Role of miRNA-15a/16 in Regulating Bmi-1 Protein in Ovarian Cancer Resistance to Cisplatin Chemotherapy
-
摘要:
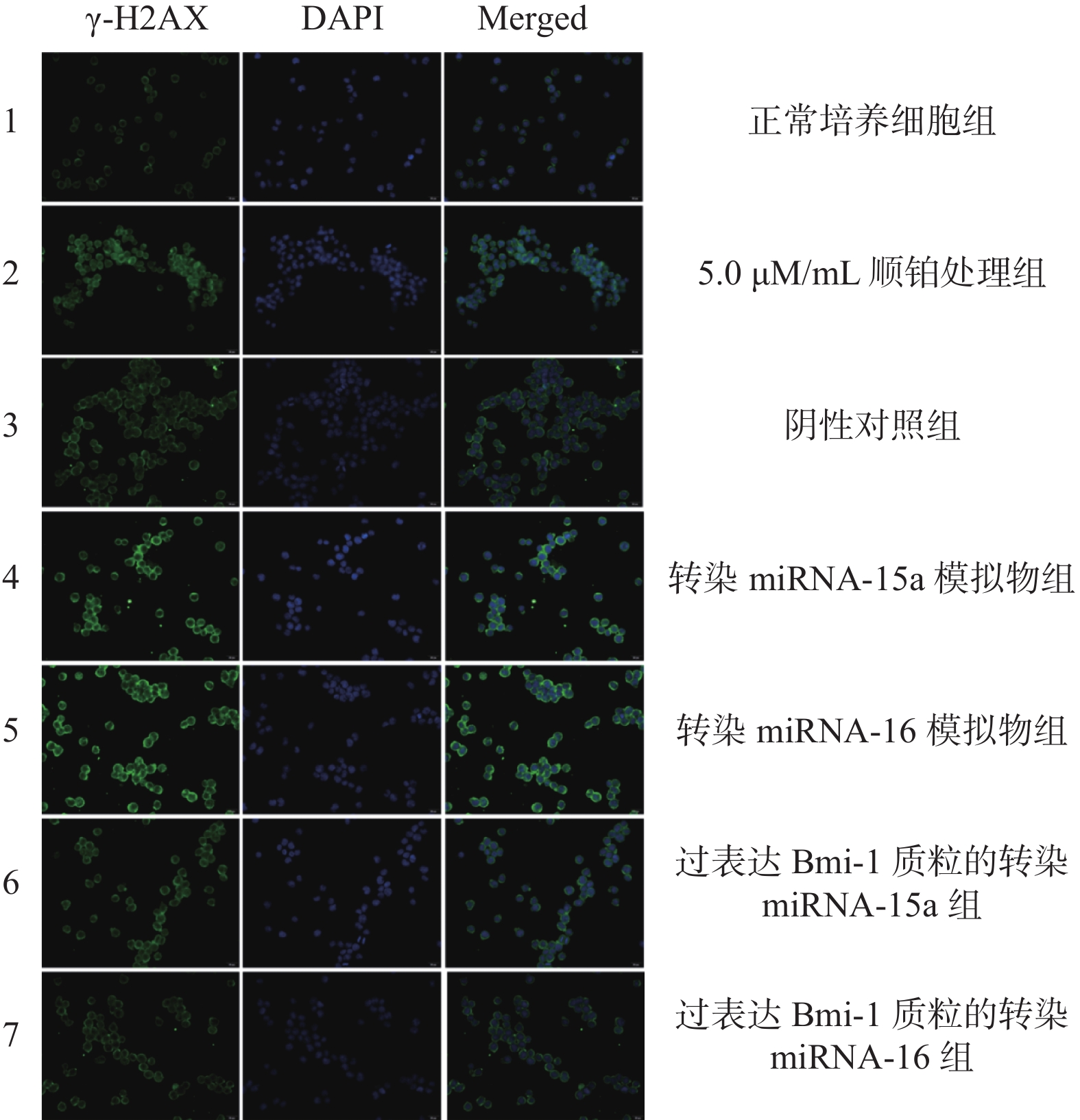
目的 探讨miRNA-15a和miRNA-16在逆转卵巢癌顺铂化疗耐药过程中的作用机制。 方法 使用miRNA-15a和miRNA-16模拟物转染人卵巢癌顺铂耐药细胞株CoC1/DDP,予顺铂处理,qRT-PCR检测正常培养CoC1/DDP细胞组、顺铂处理组、阴性对照组、转染miRNA-15a组、转染miRNA-16组和过表达的Bmi-1质粒的转染miRNA-15a和miRNA-16组的miRNA-15a、miRNA-16的表达水平,同时应用Western blot检测各组细胞中Bmi-1的表达水平,CCK-8法、Annexin V/PI法检测每组细胞的存活及凋亡情况、γ-H2AX免疫荧光检测细胞凋亡情况。 结果 卵巢癌顺铂耐药细胞株CoC1/DDP中miRNA-15a和miRNA-16低表达,Bmi-1蛋白高表达。过表达miRNA-15a、miRNA-16水平,Bmi-1蛋白下降(P < 0.05),细胞存活率下降(P < 0.05),DNA凋亡明显增加(P < 0.05),DNA损伤程度更严重(P < 0.05)。过表达Bmi-1质粒可提高细胞活度(P < 0.05),降低细胞凋亡率(P < 0.05)。 结论 Bmi-1蛋白可能是miRNA-15a和miRNA-16的调节蛋白靶点,高表达miRNA-15a和miRNA-16可以通过降低Bmi-1蛋白来提高顺铂对卵巢癌细胞的敏感性。通过对卵巢癌顺铂化疗耐药性分子标记的预测和对卵巢癌 肿耐药性目标的攻克提供了新的思路。 -
关键词:
- 卵巢癌 /
- miRNA-15a/16 /
- Bmi-1蛋白 /
- 化疗耐药 /
- 肿瘤恶性行为
Abstract:Objective To investigate the mechanisms of miRNA-15a and miRNA-16 in the process of reversing cisplatin resistance in ovarian cancer. Methods Human ovarian cancer cisplatin-resistant cell lines CoC1/DDP were transfected with miRNA-15a and miRNA-16 mimics and treated with cisplatin. qRT-PCR was used to detect the expression levels of miRNA-15a and miRNA-16 in normal CoC1/DDP cell group, cisplatin treated group, negative control group, miRNA-15a transfected group, miRNA-16 transfected group and overexpressed Bmi-1 plasmid. Western blot was used to detect the expression level of Bmi-1 in each group, CCK-8 and Annexin V/PI were used to detect cell survival and apoptosis, and γ-H2AX immunofluorescence was used to detect cell apoptosis. Results The CoC1/DDP ovarian cancer cell line shows low expression of miRNA-15a and miRNA-16, and high expression of Bmi-1 protein, which makes it resistant to cisplatin. When the levels of miRNA-15a and miRNA-16 are overexpressed, the Bmi-1 protein decreases(P < 0.05), leading to a decrease in cell survival rate(P < 0.05), a significant increase in DNA apoptosis(P < 0.05), and more severe DNA damage(P < 0.05). Overexpression of Bmi-1 plasmid can increase cell viability(P < 0.05) and reduce the rate of cell apoptosis(P < 0.05). Conclusion The Bmi-1 protein may be a target for the regulation of miRNA-15a and miRNA-16, and overexpression of miRNA-15a and miRNA-16 can increase the sensitivity of ovarian cancer cells to cisplatin by reducing the Bmi-1 protein. This provides a new idea for predicting molecular markers of cisplatin resistance in ovarian cancer and overcoming drug resistance targets in ovarian cancer. -
Key words:
- Ovarian cancer /
- miRNA-15a/16 /
- Bmi-1 protein /
- Chemotherapy resistance /
- Malignant behavior of tumor
-

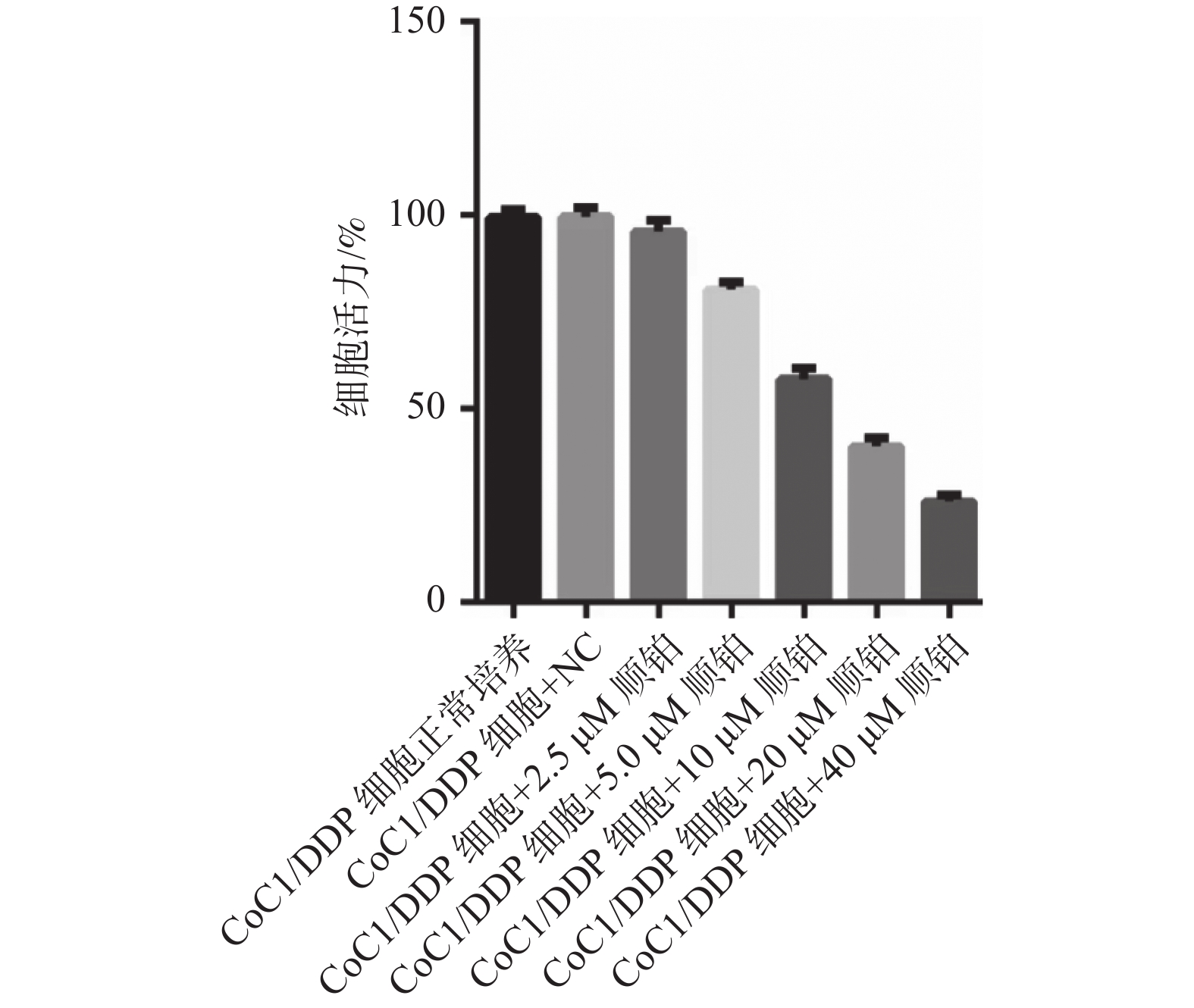
图 1 不同浓度顺铂处理的细胞活度
Figure 1. Cell activity was treated at different concentrations of cisplatin

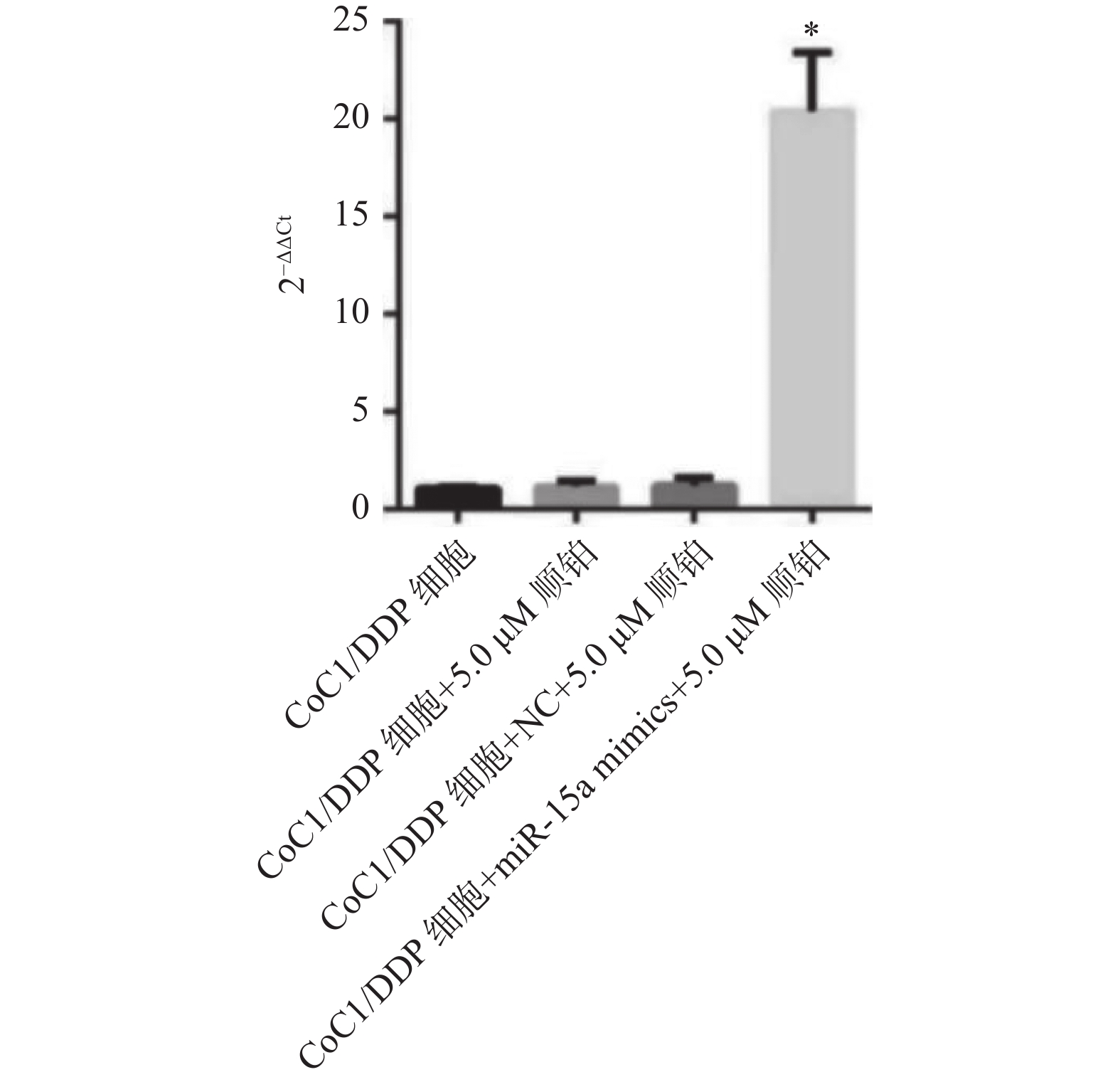
图 2 miRNA-15a在各组细胞中的表达水平
与NC组比较,*P < 0.05。
Figure 2. The expression level of miRNA15a in each group of cells

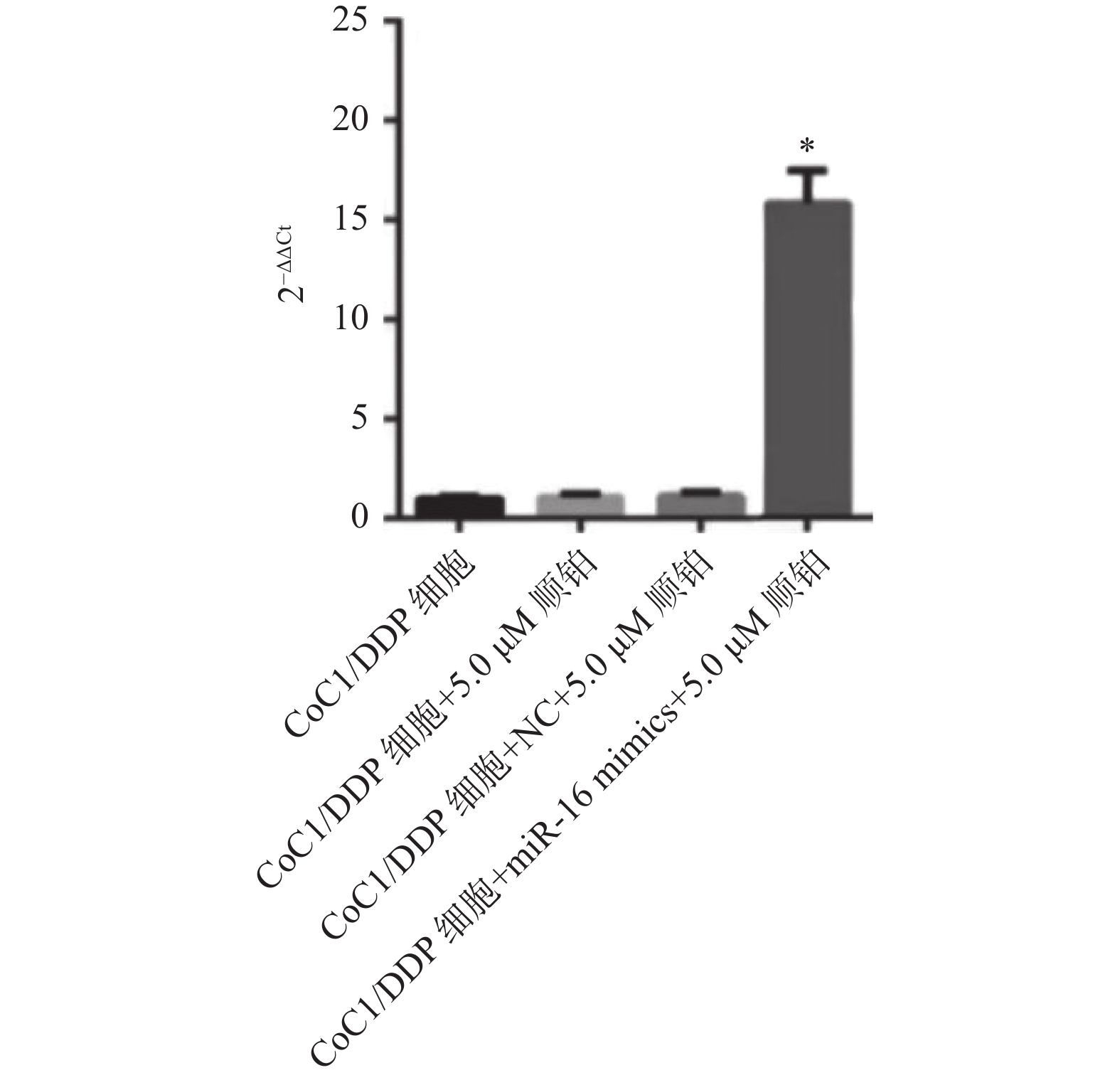
图 3 miRNA-16在各组细胞中的表达水平
与NC组比较,*P < 0.05。
Figure 3. The expression level of miRNA16 in each group of cells

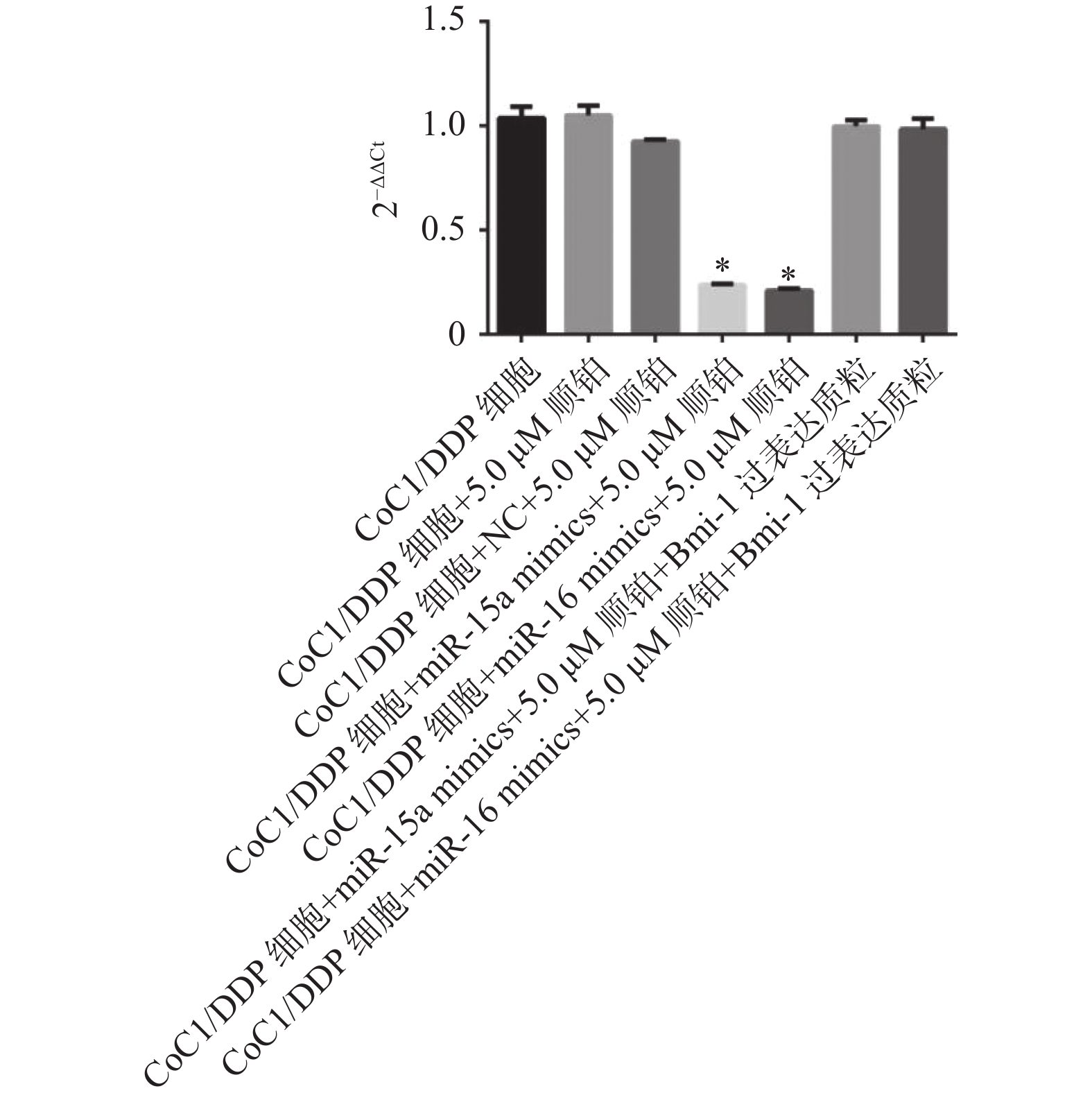
图 4 Bmi-1在各组细胞中的表达水平
与CoC1/DDP组、NC组比较,*P < 0.05。
Figure 4. The expression level of Bmi-1 in each group of cells

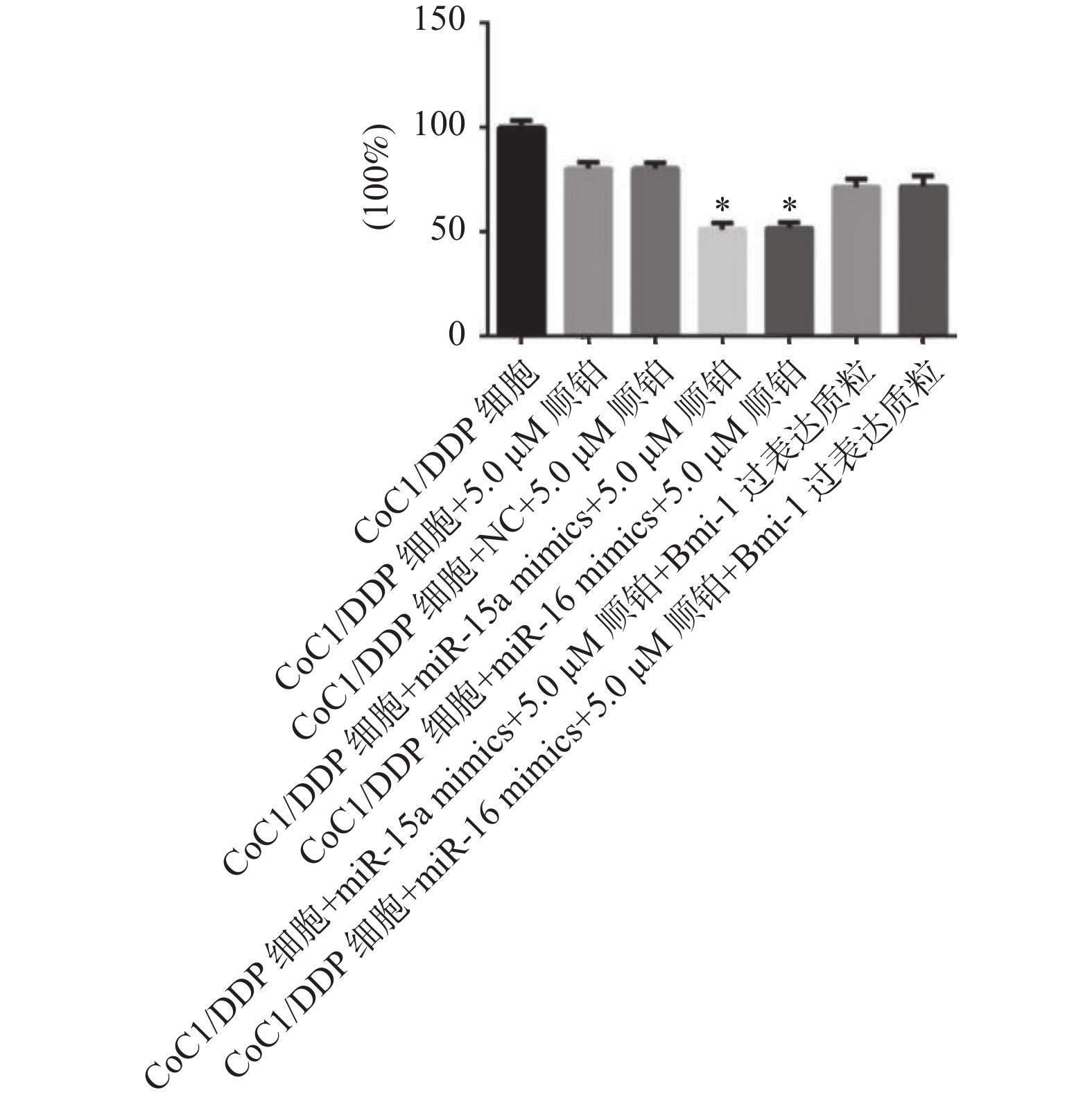
图 6 各组细胞的细胞存活度
与CoC1/DDP组、NC组、过表达质粒组比较,*P < 0.05。
Figure 6. Cell survival of each group

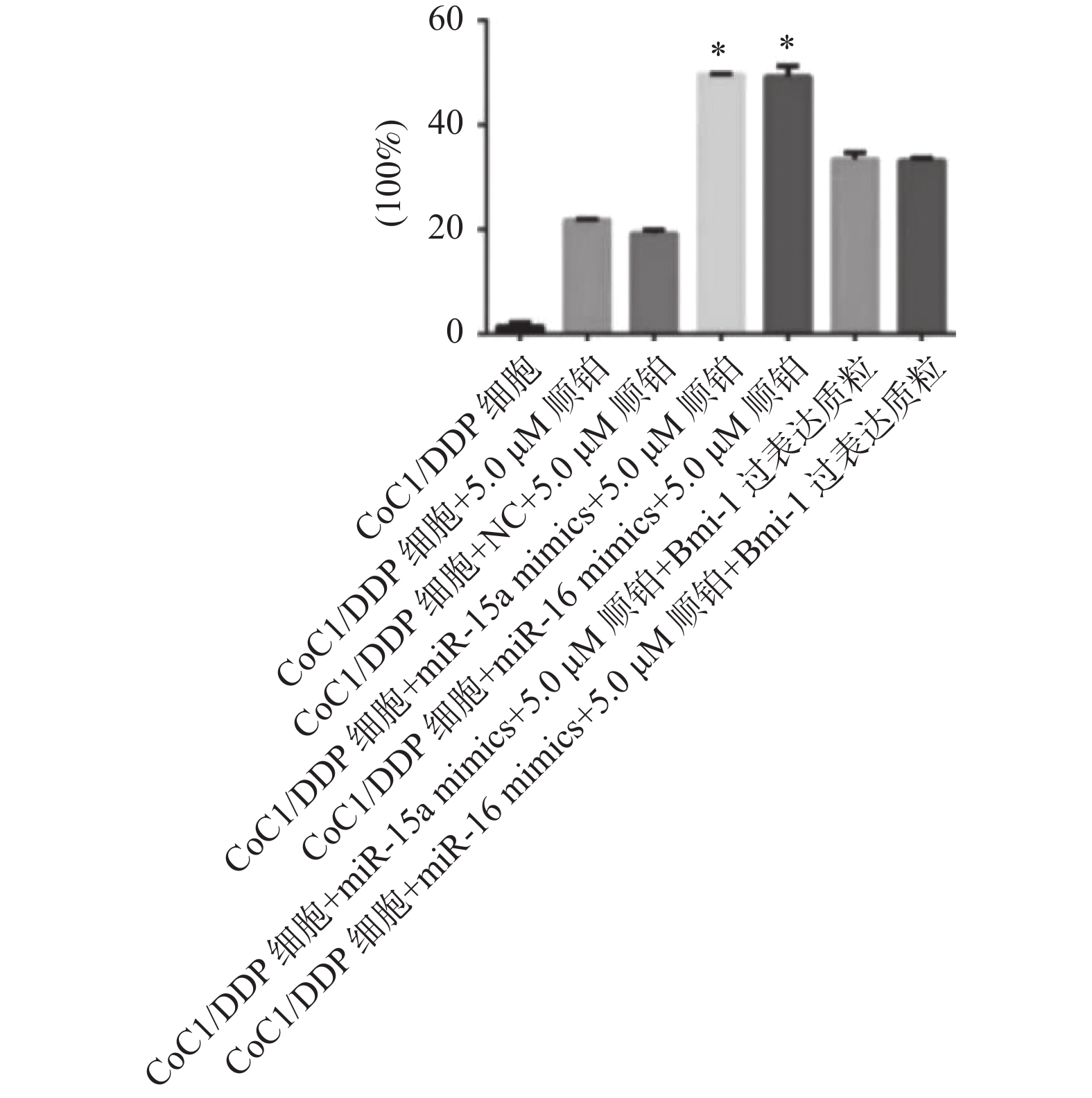
图 7 各组细胞的凋亡率水平
与CoC1/DDP组、NC组、过表达质粒组比较,*P < 0.05。
Figure 7. The apoptosis rate of cells in each group

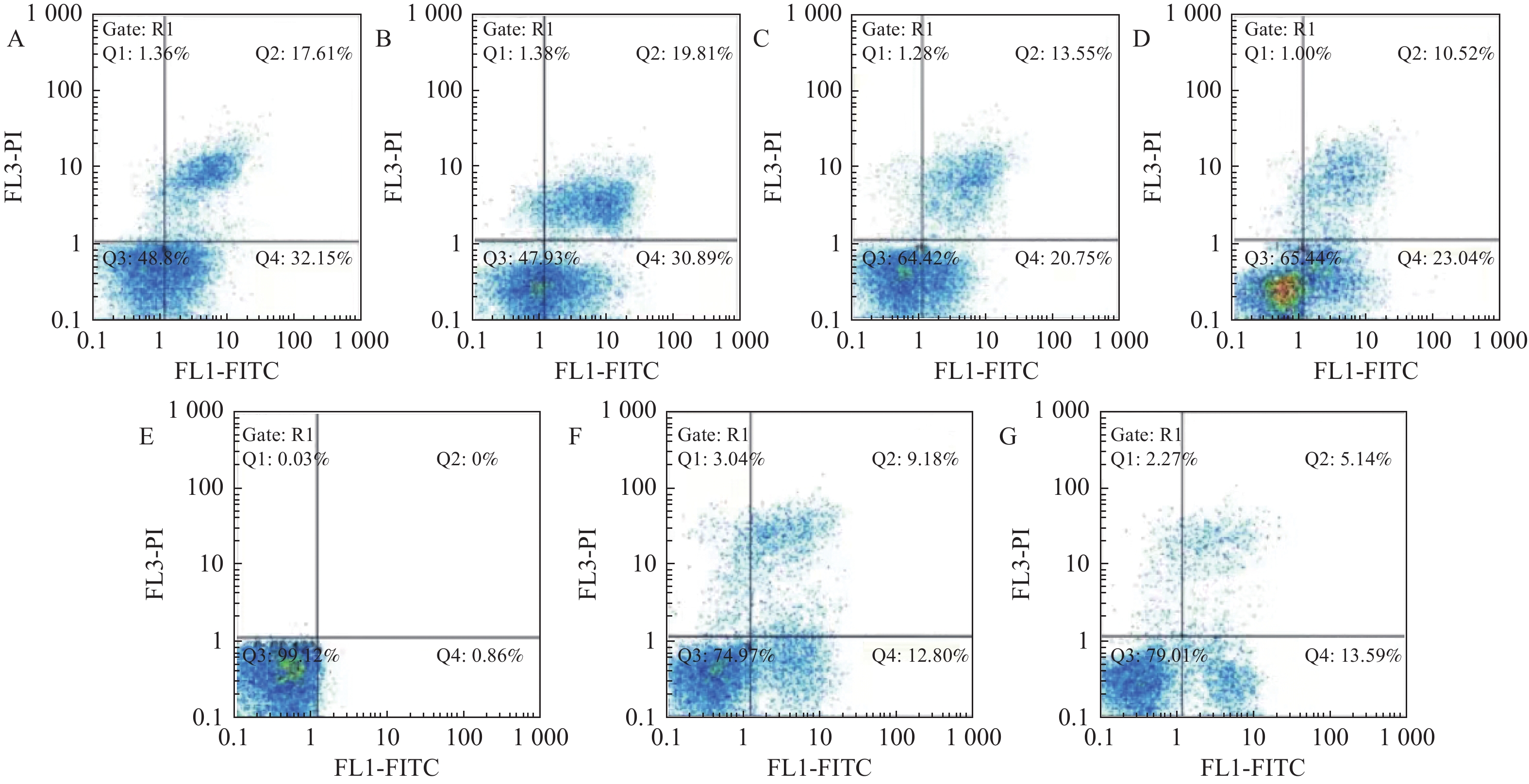
图 8 各组细胞的凋亡率图谱
A:CoC1/DDP细胞组;B:5.0 µM/mL顺铂处理组;C:NC组;D:转染miRNA-15a模拟物组;E:miRNA-16模拟物组;F:过表达的Bmi-1质粒的转染miRNA-15a组;G:过表达的Bmi-1质粒miRNA-16组。
Figure 8. Apoptotic rate atlas of each group
-
[1] Zohre M,Azita T,Safoura T,et al. Ovarian cancer in the world: Epidemiology and risk factors[J]. International Journal of Women,s Health,2019,11(1):287-299. [2] Valmiki S,Aid M A,Chaitou A R,et al. Extracellular matrix: A treasure trove in ovarian cancer dissemination and chemotherapeutic resistance[J]. Cureus,2021,13(3):e13864-13864. [3] Mylena O,Emma W,Kerry M,et al. Mechanisms of chemotherapy resistance in ovarian cancer[J]. Cancer Drug Resistance (Alhambra,Calif. ),2022,5(2):304-316. [4] Glasgow A M,Argenta P,Abrahante E J,et al. Biological insights into chemotherapy resistance in ovarian cancer[J]. International Journal of Molecular Sciences,2019,20(9):2131-2131. doi: 10.3390/ijms20092131 [5] 廉阳秧. 上皮性卵巢癌差异表达基因的生物信息学分析[D]. 昆明: 昆明医科大学, 2016. [6] Wu C,Zheng X,Li X,et al. Reduction of gastric cancer proliferation and invasion by miR-15a mediated suppression of Bmi-1 translation[J]. Oncotarget,2016,7(12):14522-14536. doi: 10.18632/oncotarget.7392 [7] Penny S M. Ovarian cancer: An overview[J]. Radiol Technol,2020,91(6):561-575. [8] Liu Q,Novak M K,Pepin R M,et al. MicroRNA-mediated regulation of microRNA machinery controls cell fate decisions[J]. ELife,2021,10:e72289. doi: 10.7554/eLife.72289 [9] Kabakov A V,Kazakov O V,Poveshchenko A F,et al. Correlation between structural transformations in mesenteric lymph nodes and the levels microRNA during polychemotherapy of breast cancer[J]. Bulletin of Experimental Biology and Medicine,2022,172(4):467-471. doi: 10.1007/s10517-022-05415-4 [10] 杨雷,魏丽军,王福花,等. miR-93-3p和CDC42在上皮性卵巢癌化疗耐药组织中的表达及临床意义[J]. 实用医学杂志,2020,36(9):1217-1222. doi: 10.3969/j.issn.1006-5725.2020.09.020 [11] Kulkarni B,Kirave P,Gondaliya P,et al. Exosomal miRNA in chemoresistance,immune evasion,metastasis and progression of cancer[J]. Drug Discovery Today,2019,24(10):2058-2067. doi: 10.1016/j.drudis.2019.06.010 [12] Bugra S T,Demet A,Betul C,et al. The expression levels of miRNA-15a and miRNA-16-1 in circulating tumor cells of patients with diffuse large B-cell lymphoma[J]. Molecular Biology Reports,2018,46(1):975-980. [13] Xu,P,Wang Y,Deng Z,et al. MicroRNA-15a promotes prostate cancer cell ferroptosis by inhibiting GPX4 expression[J]. Oncology Letters,2022,23(2):67. doi: 10.3892/ol.2022.13186 [14] Chu J,Zhu Y,Liu Y,et al. E2F7 overexpression leads to tamoxifen resistance in breast cancer cells by competing with E2F1 at miR-15a/16 promoter[J]. Oncotarget,2015,6(31):31944-31957. doi: 10.18632/oncotarget.5128 [15] Dhar K S D,Banerjee S M,Dahai J,et al. Therapeutic evaluation of microRNA-15a and microRNA-16 in ovarian cancer[J]. Oncotarget,2016,7(12):15093-15104. doi: 10.18632/oncotarget.7618 [16] Dey A,Xiong X,Crim A,et al. Evaluating the mechanism and therapeutic potential of PTC-028,a novel inhibitor of BMI-1 function in ovarian cancer[J]. Molecular Cancer Therapeutics,2018,17(1):39-49. doi: 10.1158/1535-7163.MCT-17-0574 [17] Xin T,Zhang F B,Sui G J,et al. Bmi-1 siRNA inhibited ovarian cancer cell line growth and decreased telomerase activity[J]. British Journal of Biomedical Science,2012,69(2):62-66. doi: 10.1080/09674845.2012.12002438 [18] Zhang X L,Sun B L,Tian S X,et al. MicroRNA-132 reverses cisplatin resistance and metastasis in ovarian cancer by the targeted regulation on Bmi-1[J]. European Review for Medical and Pharmacological Sciences,2019,23(9):3635-3644. [19] Salem A,Hagen K,Silvia D,et al. Polycomb protein BMI-1 as a potential therapeutic target in mucinous ovarian cancer[J]. Anticancer Research,2022,42(4):1739-1747. doi: 10.21873/anticanres.15650 -

 下载:
下载:

 点击查看大图
点击查看大图
计量
- 文章访问数: 1364
- HTML全文浏览量: 847
- PDF下载量: 13
- 被引次数: 0
