

Practice and Exploration of the Cultivation of Scientific Innovation Ability Based on RBL Teaching Method
-
摘要:
目的 探索基于研究的学习(research based learning,RBL)教学法对本科生科研创新能力的影响。 方法 一共212 名本科生纳入研究,其中传统教学组有104名,RBL教学组有108名,之后,教师对2组学生的科研创新能力进行客观评价,并调查了学生对教学效果的满意度。 结果 RBL教学组本科生的科研创新能力总分高于传统教学组(P < 0.001),RBL教学组本科生在实验设计、实验操作、报告撰写、结果汇报、提问问题与回答问题能力,以及对教学效果的满意度均高于传统教学组(P < 0.01)。 结论 RBL 教学法可以提高本科生的科研创新能力。 Abstract:Objective To explore the impact of research based learning(RBL) teaching method on undergraduate research innovation ability. Methods A total of 212 undergraduate students were involved in this study. Among them, 104 undergraduate students participated in the traditional teaching method, and 108 undergraduate students participated in the RBL teaching method. After they finished the comprehensive experiment in medical immunology, we assessed the scientific innovation ability and the degree of satisfaction of teaching effectiveness. Results The undergraduate research and innovation capabilities of the RBL teaching group are higher than those of the traditional teaching group(P < 0.001). The undergraduate students in the RBL teaching group have higher abilities in experimental design, experimental operation, report writing, result presentation, questioning and answering, as well as satisfaction with teaching effectiveness, compared to the traditional teaching group(P < 0.01). Conclusion The RBL teaching method can significantly improve the scientific innovation ability of undergraduate students. -

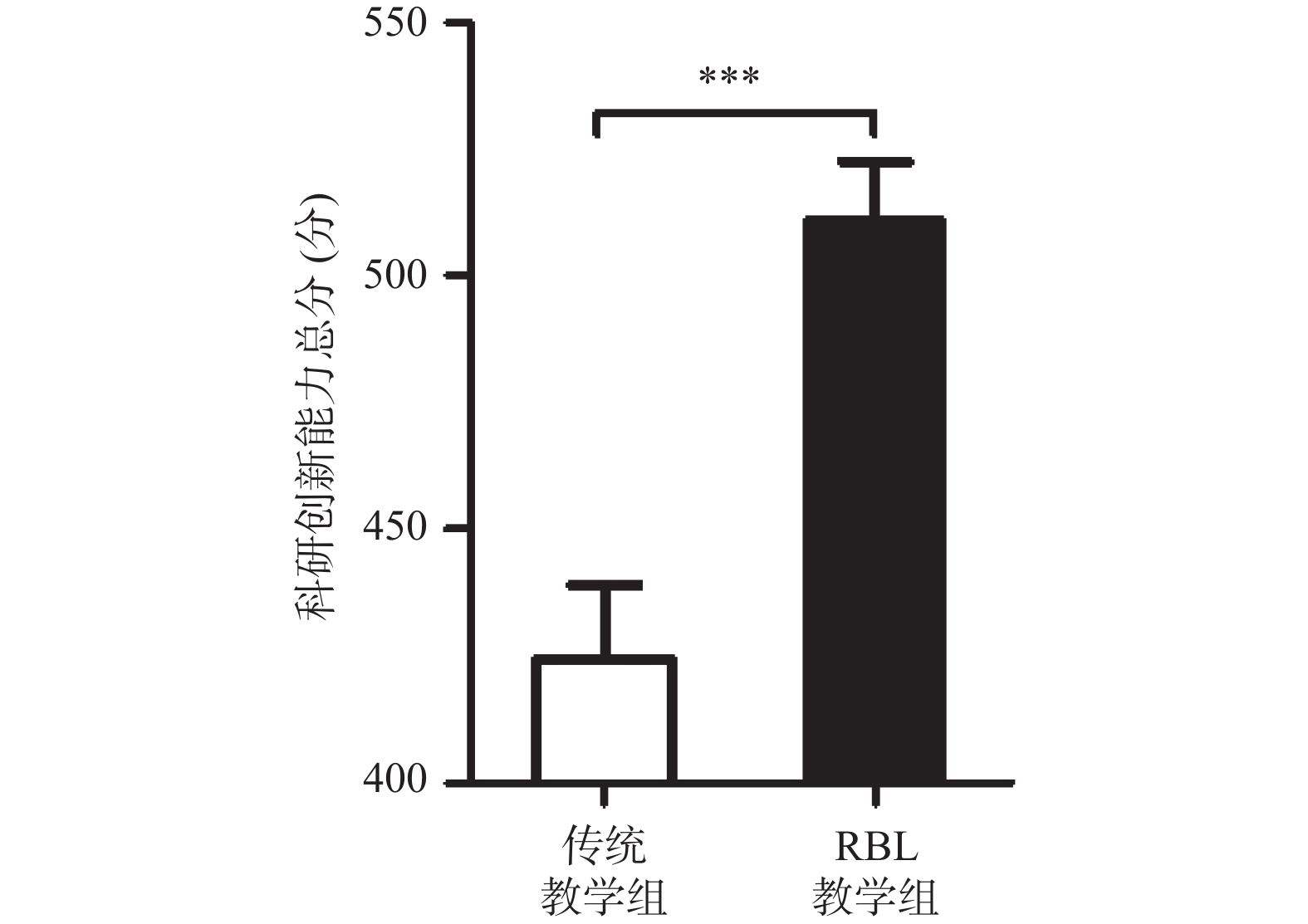
图 1 传统教学组和RBL教学组学生科研创新能力总分得分比较
***P < 0.001。
Figure 1. Comparison of the total score of the scientific innovation ability between traditional teaching group and RBL teaching group

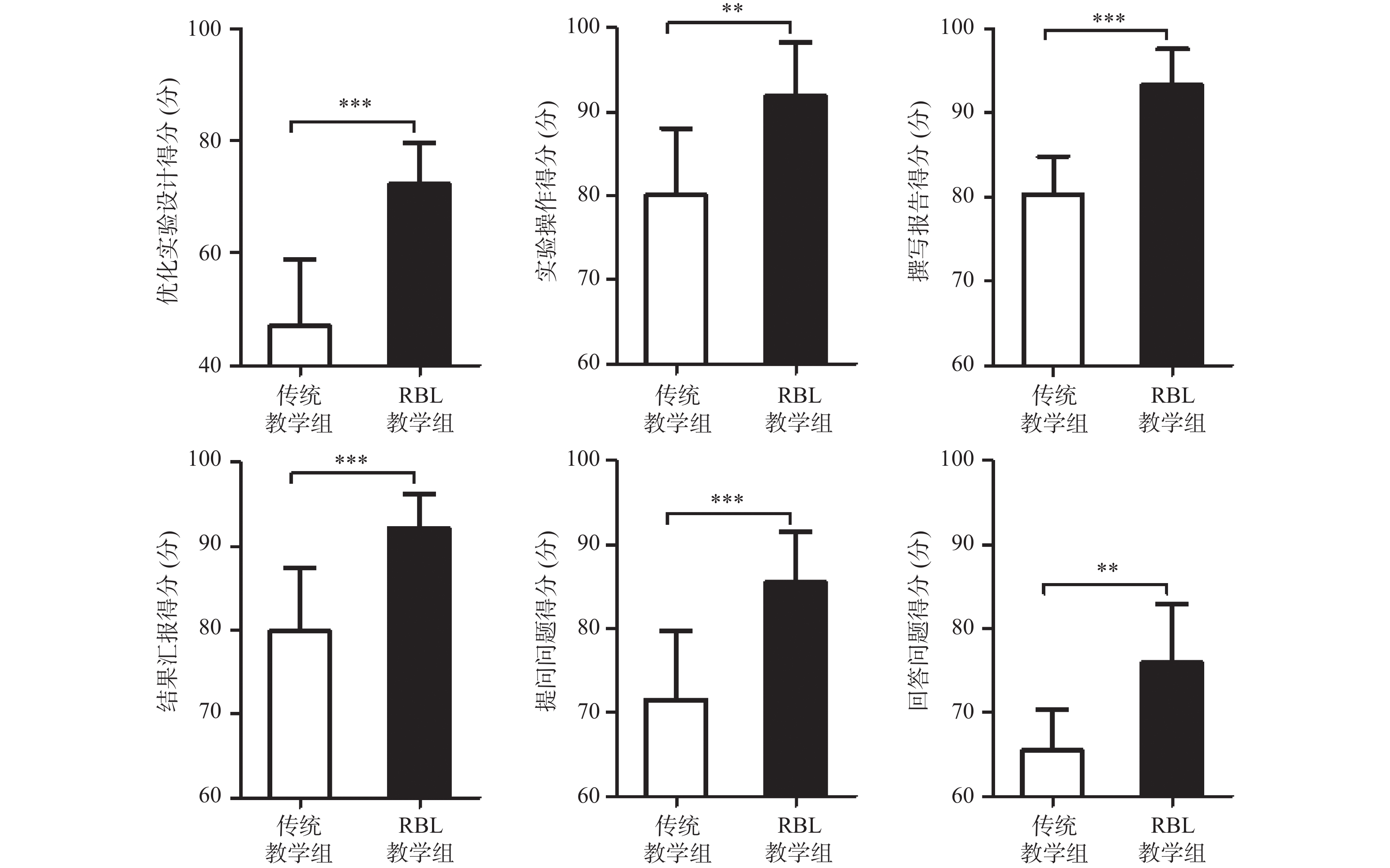
图 2 传统教学组和RBL教学组学生科研创新能力各维度得分比较
**P < 0.01,***P < 0.001。
Figure 2. Comparison of the scores of each dimension between traditional teaching group and RBL teaching group

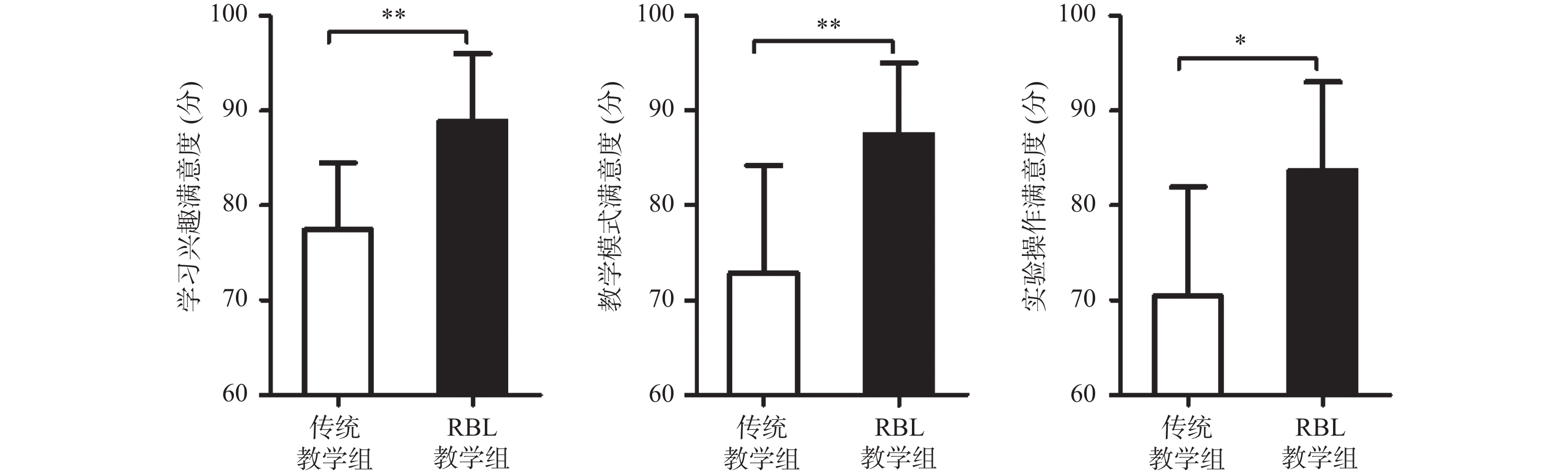
图 3 传统教学组和RBL教学组学生对教学满意度的评分比较
*P < 0.05,***P < 0.01。
Figure 3. Comparison of the scores of degree of satisfaction between traditional teaching group and RBL teaching group
表 1 传统教学组和RBL教学组学生基本资料比较($ \bar x \pm s $)
Table 1. Comparison of the basic information between the traditional teaching group and RBL teaching group ($ \bar x \pm s $)
组别 学生
(n)男生
(n)女生
(n)平均
年龄(岁)免疫学
实验基础GPA 传统教学组 104 50 54 21.16 ± 1.16 无 3.52 ± 0.39 RBL教学组 108 53 55 20.89 ± 1.07 无 3.47 ± 0.43 F — — — 1.285 — 1.082 P — — — 0.273 — 0.311  下载: 导出CSV
下载: 导出CSV
-
[1] 许日华,乐传永. 新型研究型大学战略空间构造与创新[J]. 教育研究,2023,44(7):81-93. [2] 吕莹,陈璧州. 教学研究型大学学生科研能力培养探析[J]. 教育教学论坛,2014(3):69-71. [3] 冯蕊,徐世莲,杨海龙,等. RBL教学法对医学研究生批判性思维的影响[J]. 昆明医科大学学报,2023,44(10):196-202. doi: 10.12259/j.issn.2095-610X.S20231008 [4] Li C. English research learning and functional research based on constructivism theory and few-shot learning[J]. Computational Intelligence And Neuroscience,2022,2022:3698802. [5] Rosario R,Hopper T S,Huang-Saad A. Applying research-based teaching strategies in a biomedical engineering programming course: Introduction to computer aided diagnosis[J]. Biomeical Engineering Education,2022,2(1):41-59. doi: 10.1007/s43683-021-00057-w [6] Benning N,Knaup P. Development of a research-based teaching course as blended-learning format in a medical informatics program[J]. Studies in Health Technology and Informatics,2019,264:1909-1910. [7] 邓颖,黄聪,张楠楠,等. 基于RBL教学模式在药理学教学中进行生物信息与多组学教学的探索[J]. 贵州中医药大学学报,2023,45(04):87-90. [8] Wesselborg B,Hoenen M,Adam-Paffrath R,et al. Interprofessional nutrition management-implementation and evaluation of a course for medical and nursing students using research-based learning method[J]. GMS Journal for Medical Education,2019,36(6):68. [9] 王刚,张秋香,毛丙永,等. 本科生科研兴趣与能力培养的探索与实践[J]. 科教文汇,2020(34):26-27. [10] Holmes N,Wieman C,Bonn D. Teaching critical thinking[J]. Proceedings of the National Academy of Sciences of the United States of America,2015,112(36):11199-11204. [11] 付坤,王瑞,杨罕,等. 高校本科生科研素养培养教育探索[J]. 实验室研究与探索,2017,36(3):207-211. [12] 毛文,刘清心. 以学生为中心的设计方法课程多元化教学模式探索[J]. 河南教育(高等教育),2023 , (7):93-94. [13] Carter A,Creedy D,Sidebotham M. Efficacy of teaching methods used to develop critical thinking in nursing and midwifery undergraduate students: A systematic review of the literature[J]. Nurse Education Today,2016,40:209-218. doi: 10.1016/j.nedt.2016.03.010 [14] Granjeiro É. Research-based teaching-learning method: A strategy to motivate and engage students in human physiology classes[J]. Advances in Physiology Education,2019,43(4):553-556. doi: 10.1152/advan.00034.2019 [15] 徐志鹏,邱竞帆,王云峰. 在强化医学文献阅读中提升医学生的科学素养和科研创新能力[J]. 南京医科大学学报(社会科学版),2021,21(2):189-192. [16] 钮晓音,郭晓奎. “新医科”背景下的医学教育改革与人才培养[J]. 中国高等医学教育,2021 , (5):1-2. [17] 陈戬,赵耘佩,吴雨菲. 医学本科生创新能力的培养策略研究[J]. 医学研究杂志,2021,50(9):163-165. [18] 张伟,祁俊侠,杨茜璐,等. 基于文献阅读和科研设计培养医学本科生的科研素养[J]. 基础医学教育,2023,25(3):261-265. [19] Ghukasyan G,Kurghinyan M,Hovhannisyan L,et al. Moving research-based learning in life sciences upstream,and beyond borders: An international group research project for high school youth[J]. Omics:A Journal of Integrative Biology,2023,27(9):421-425. doi: 10.1089/omi.2023.0079 -

 点击查看大图
点击查看大图
计量
- 文章访问数: 1752
- HTML全文浏览量: 1212
- PDF下载量: 28
- 被引次数: 0
