

Culture of Malignant Pleural Mesothelioma Cells and the Effects of CDKN2B on Cancer Cell
-
摘要:
目的 探讨不同培养条件(RPMI-1640、DMEM和DMEM/F12培养液)对人恶性胸膜间皮瘤(malignant pleural mesothelioma,MPM)组织中分离的MPM细胞传代的影响以及细胞周期蛋白依赖性激酶抑制剂2B(cyclin dependent kinase inhibitor 2B,CDKN2B)对MPM细胞增殖、侵袭和凋亡的作用。 方法 从MPM组织中分离细胞分别用RPMI-1640、DMEM和DMEM/F12培养液培养。CCK-8检测细胞增殖,细胞核及染色体利用瑞氏-吉姆萨染色观察,免疫荧光实验检测MPM标志物Calretinin、CD141、CK5、EMA和WT-1荧光强度。RT-qPCR和Western blot分别检测CDKN2B的mRNA和蛋白表达能力。Transwell检测细胞侵袭能力,流式细胞术检测细胞凋亡率。 结果 建立的MPM细胞在RPMI-1640、DMEM和DMEM/F12培养液中传至第10代仍具有较好的活力,且MPM标志物Calretinin、CD141、CK5、EMA和WT-1在细胞中均表达,MPM细胞在RPMI-1640培养液中活力较为稳定。CDKN2B在MPM细胞中低表达(P < 0.05),过表达CDKN2B显著抑制MPM细胞的增殖(P < 0.05)、侵袭(P < 0.05)和上皮间质转化(P < 0.01),促进细胞凋亡(P < 0.01)。 结论 建立的MPM细胞可在RPMI-1640培养液中稳定传代,CDKN2B可作为MPM诊断和治疗的潜在靶标。 Abstract:Objective To investigate the effects of different culture conditions(RPMI-1640, DMEM and DMEM/F12 medium) on the passage of MPM cells isolated from the tissues of Malignant pleural mesothelioma(MPM), and to study the effects of CDKN2B on the proliferation, invasion and apoptosis of MPM cells. Methods MPM cells were isolated from MPM tissues and cultured in RPMI-1640, DMEM and DMEM/F12 medium, respectively. Cell proliferation was examined by CCK-8, and the nuclei and chromosomes were observed by Wright-Giemsa staining. Fluorescence intensities of Calretinin, CD141, CK5, EMA and WT-1 were conducted by immunofluorescence assay. The mRNA and protein expression of CDKN2B were detected by RT-qPCR and Western blot, respectively. Transwell was used to detect cell invasion and flow cytometry was used to detect cell apoptosis. Results The established MPM cells showed good viability when passaged to the 10th generation in RPMI-1640, DMEM and DMEM/F12 cultures, and the MPM markers Calretinin, CD141, CK5, EMA and WT-1 were all expressed in the cells. The viability of MPM cells in RPMI-1640 culture medium was relatively stable. CDKN2B was downregulated in MPM cells(P < 0.05), and overexpression of CDKN2B significantly suppressed the proliferation(P < 0.05), invasion(P < 0.05) and epithelial interstitial transformation of MPM cells(P < 0.01), and promoted the apoptosis(P < 0.01). Conclusion The established MPM cells were stably passaged in RPMI-1640 culture medium, and CDKN2B may be a potential target for the diagnosis and treatment of MPM. -
Key words:
- Malignant pleural mesothelioma /
- Culture conditions /
- RPMI-1640 culture medium /
- CDKN2B /
- Proliferation /
- Invasion
-

图 1 3种培养条件对MPM细胞形态和增殖的作用
A:倒置显微镜下观察P1、P5和P10细胞形态(100×);B:CCK-8检测P1、P5和P10细胞细胞增殖活力;与RPMI-1640组比较,*P < 0.05;与DMEM组比较,△P < 0.05;P1:第1代;P5:第5代;P10:第10代。
Figure 1. Effect of three culture conditions on morphology and proliferation of MPM cells

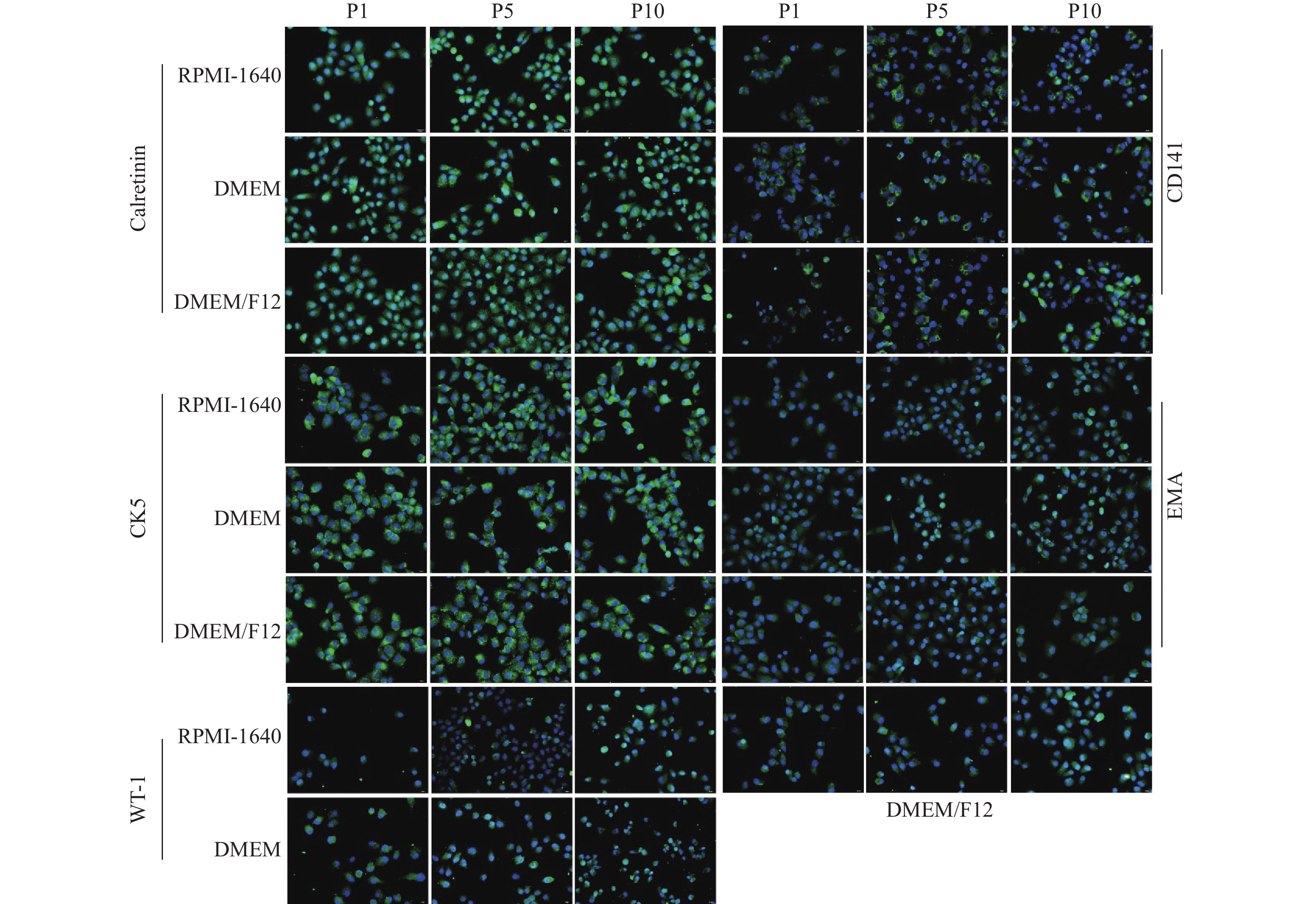
图 3 免疫荧光检测MPM细胞标志物(40×)
Figure 3. The MPM cells biomarkers were detected by immunofluorescence assay(40×)

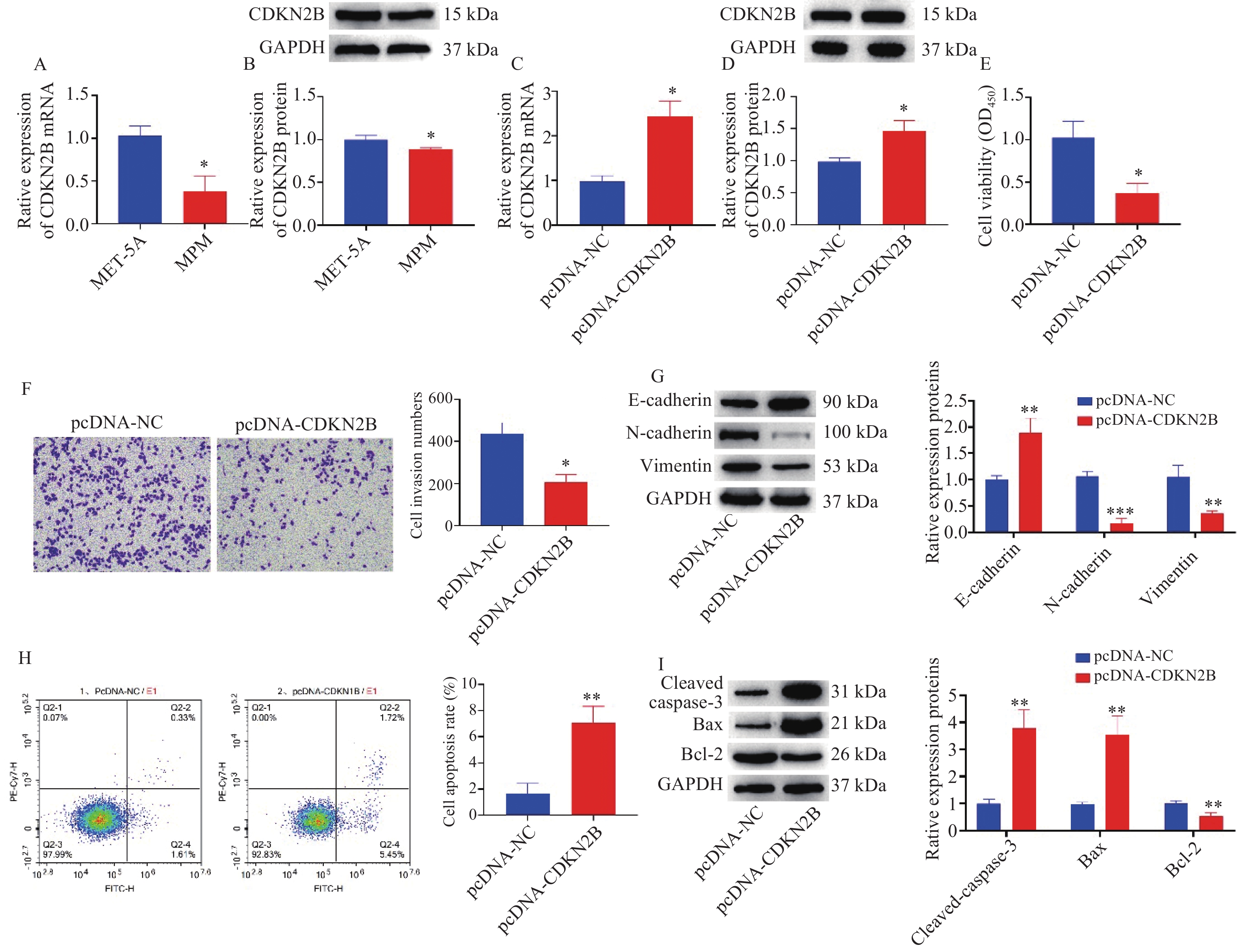
图 4 CDKN2B调控MPM细胞增殖、侵袭、上皮间质转化和凋亡
A:RT-qPCR检测CDKN2B mRNA表达;B:Western blot检测CDKN2B蛋白表达;与MET-5A组相比,*P < 0.05;C:RT-qPCR检测pcDNA-CDKN2B转染效率;D:Western blot检测pcDNA-CDKN2B转染效率;E:CCK-8检测细胞增殖活力;F:Transwell检测细胞侵袭能力;G:Western blot检测细胞上皮间质转化相关标志物的蛋白表达;H:流式细胞术检测细胞凋亡率;I:Western blot检测细胞凋亡相关标志物的蛋白表达;与pcDNA-NC组比较,*P < 0.05,**P < 0.01,***P < 0.001。
Figure 4. CDKN2B modulated the proliferation,invasion,epithelial interstitial transformation and apoptosis of MPM cells
-
[1] Bruno R,Alì G,Fontanini G. Molecular markers and new diagnostic methods to differentiate malignant from benign mesothelial pleural proliferations: A literature review[J]. J Thorac Dis,2018,10(Suppl 2):S342-S352. [2] Stumphius J,Meyer P B. Asbestos bodies and mesothelioma[J]. Ann Occup Hyg,1968,11(4):283-293. [3] Porret E, Madelaine J, Galateau-Sallé F, et al. Epidemiology, molecular biology, diagnostic and therapeutic strategy of malignant pleural mesothelioma in 2007 - an update[J]. Rev Mal Respir, 2007, 24(8 Pt 2): 6S157-6S164. [4] Robinson B W,Musk A W,Lake R A. Malignant mesothelioma[J]. Lancet,2005,366(9483):397-408. doi: 10.1016/S0140-6736(05)67025-0 [5] Santoro A,O'brien M E,Stahel R A,et al. Pemetrexed plus cisplatin or pemetrexed plus carboplatin for chemonaïve patients with malignant pleural mesothelioma: Results of the international expanded access program[J]. J Thorac Oncol,2008,3(7):756-763. doi: 10.1097/JTO.0b013e31817c73d6 [6] Macleod N,Chalmers A,O'rourke N,et al. Is radiotherapy useful for treating pain in mesothelioma? A phase ii trial[J]. J Thorac Oncol,2015,10(6):944-950. doi: 10.1097/JTO.0000000000000499 [7] Clive A O,Taylor H,Dobson L,et al. Prophylactic radiotherapy for the prevention of procedure-tract metastases after surgical and large-bore pleural procedures in malignant pleural mesothelioma (SMART): A multicentre,open-label,phase 3,randomised controlled trial[J]. Lancet Oncol,2016,17(8):1094-1104. doi: 10.1016/S1470-2045(16)30095-X [8] Zalcman G,Mazieres J,Margery J,et al. Bevacizumab for newly diagnosed pleural mesothelioma in the mesothelioma avastin cisplatin pemetrexed study (MAPS): A randomised,controlled,open-label,phase 3 trial[J]. Lancet,2016,387(10026):1405-1414. doi: 10.1016/S0140-6736(15)01238-6 [9] Chernova T,Sun X M,Powley I R,et al. Molecular profiling reveals primary mesothelioma cell lines recapitulate human disease[J]. Cell Death Differ,2016,23(7):1152-1164. doi: 10.1038/cdd.2015.165 [10] Manning L S,Whitaker D,Murch A R,et al. Establishment and characterization of five human malignant mesothelioma cell lines derived from pleural effusions[J]. Int J Cancer,1991,47(2):285-290. doi: 10.1002/ijc.2910470219 [11] Kobayashi M,Takeuchi T,Ohtsuki Y. Establishment of three novel human malignant pleural mesothelioma cell lines: Morphological and cytogenetical studies and EGFR mutation status[J]. Anticancer Res,2008,28(1a):197-208. [12] Philippeaux M M,Pache J C,Dahoun S,et al. Establishment of permanent cell lines purified from human mesothelioma: Morphological aspects,new marker expression and karyotypic analysis[J]. Histochem Cell Biol,2004,122(3):249-260. doi: 10.1007/s00418-004-0701-1 [13] Kanellakis N I,Asciak R,Hamid M A,et al. Patient-derived malignant pleural mesothelioma cell cultures: A tool to advance biomarker-driven treatments[J]. Thorax,2020,75(11):1004-1008. doi: 10.1136/thoraxjnl-2020-215027 [14] Oey H,Daniels M,Relan V,et al. Whole-genome sequencing of human malignant mesothelioma tumours and cell lines[J]. Carcinogenesis,2019,40(6):724-734. doi: 10.1093/carcin/bgz066 [15] Szulkin A,Nilsonne G,Mundt F,et al. Variation in drug sensitivity of malignant mesothelioma cell lines with substantial effects of selenite and bortezomib, highlights need for individualized therapy[J]. PLoS One,2013,8(6):e65903. [16] Yuile A,Satgunaseelan L,Wei J Q,et al. CDKN2A/B homozygous deletions in astrocytomas: A literature review[J]. Curr Issues Mol Biol,2023,45(7):5276-5292. doi: 10.3390/cimb45070335 [17] Pozdeyev N,Gay L M,Sokol E S,et al. Genetic analysis of 779 advanced differentiated and anaplastic thyroid cancers[J]. Clin Cancer Res,2018,24(13):3059-3068. [18] Fortin Ensign S P,Jenkins R B,Giannini C,et al. Translational significance of CDKN2A/B homozygous deletion in isocitrate dehydrogenase-mutant astrocytoma[J]. Neuro Oncol,2023,25(1):28-36. doi: 10.1093/neuonc/noac205 [19] Yang L,Ma D W,Cao Y P,et al. PRMT5 functionally associates with EZH2 to promote colorectal cancer progression through epigenetically repressing CDKN2B expression[J]. Theranostics,2021,11(8):3742-3759. doi: 10.7150/thno.53023 [20] Yang J,Liu Y,He A,et al. Hsa-miR-429 promotes bladder cancer cell proliferation via inhibiting CDKN2B[J]. Oncotarget,2017,8(40):68721-68729. doi: 10.18632/oncotarget.19878 [21] Cao K,Li B,Zhang Y W,et al. miR-29b restrains cholangiocarcinoma progression by relieving DNMT3B-mediated repression of CDKN2B expression[J]. Aging (Albany NY),2021,13(4):6055-6065. doi: 10.18632/aging.202549 [22] Boldrin E,Gaffo E,Niedermayer A,et al. MicroRNA-497/195 is tumor suppressive and cooperates with CDKN2A/B in pediatric acute lymphoblastic leukemia[J]. Blood,2021,138(20):1953-1965. doi: 10.1182/blood.2020007591 -

 下载:
下载:

 点击查看大图
点击查看大图

计量
- 文章访问数: 1678
- HTML全文浏览量: 937
- PDF下载量: 20
- 被引次数: 0
