

Expression of PDE4D Gene in Ovarian Cancer Based on Oncomine and the Regulation of Sanguinarine
-
摘要:
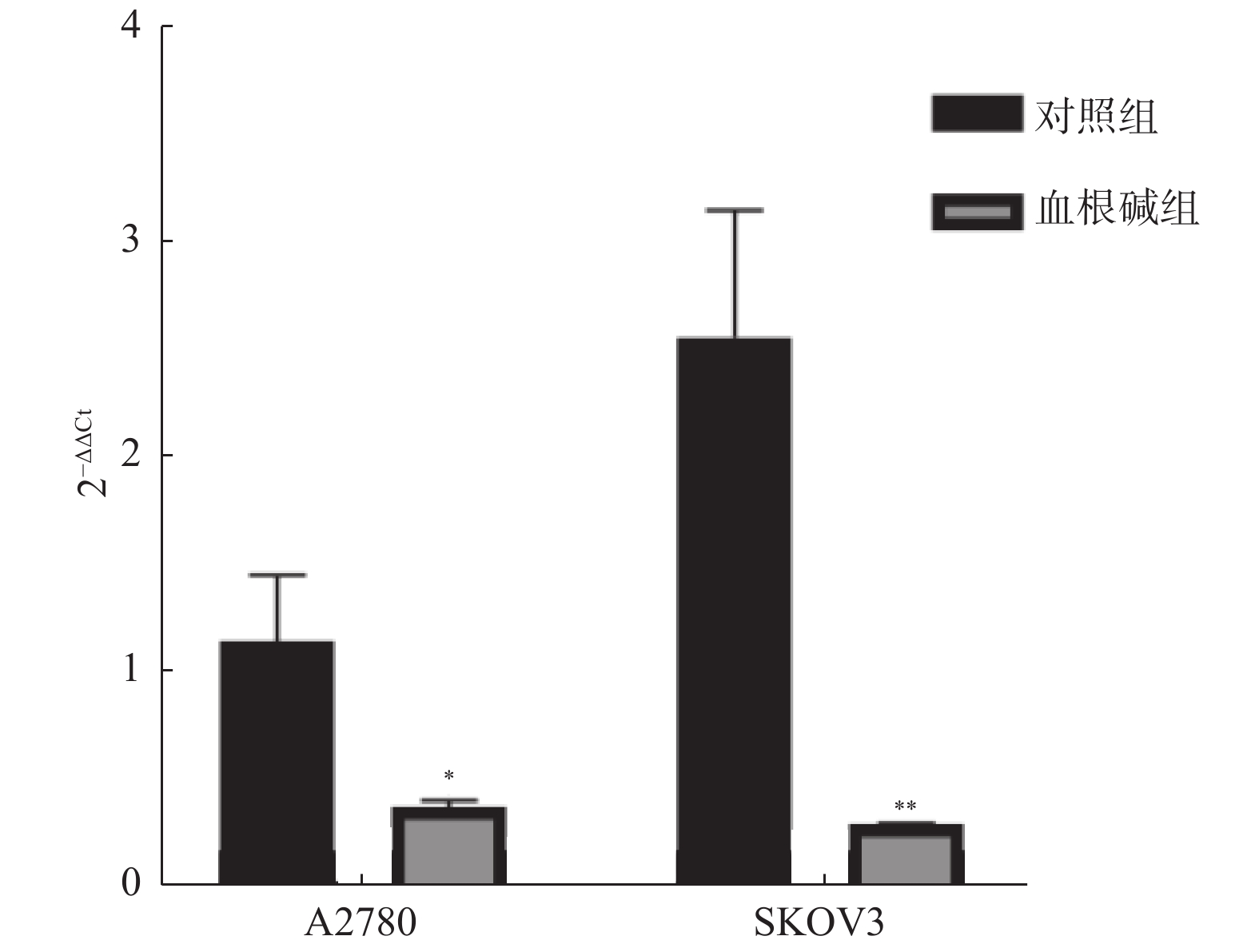
目的 通过深度挖掘Oncomine数据库中PDE4D在卵巢癌的研究数据,分析PDE4D基因在卵巢癌中的表达变化,并进行细胞实验研究血根碱对卵巢癌细胞中PDE4D基因表达的影响。 方法 PharmMapper数据库查询与血根碱药效基团匹配的作用靶点,收集Oncomine数据库 PDE4D在卵巢癌中的研究数据,进一步分析PDE4D在卵巢癌中的表达变化。将卵巢癌SKOV3和A2780细胞分成对照组和血根碱组,CCK8和RT-qPCR法分别检测血根碱对卵巢癌A2780、SKOV3细胞增殖及PDE4D mRNA表达的影响。 结果 PDE4D是与血根碱药效基团匹配的作用靶点。Oncomine数据库中共查询到6项正常卵巢组织和卵巢癌组织PDE4D基因的研究,且PDE4D在不同类型的卵巢癌组织中均呈高表达(P < 0.05)。细胞实验表明,与对照组相比,血根碱组卵巢癌A2780、SKOV3细胞增殖被明显抑制,PDE4D mRNA表达降低,差异均有统计学意义(P < 0.05)。 结论 卵巢癌组织中PDE4D基因呈高表达,血根碱抑制卵巢癌细胞生长可能与下调PDE4D表达相关。 -
关键词:
- 卵巢癌 /
- PDE4D /
- Oncomine数据库 /
- 血根碱
Abstract:Objective To investigate the expression and clinical significance of PDE4D in ovarian cancer by excavating gene information in Oncomine and explore the effect of sanguinarine on PDE4D by cells experiment. Methods PharmMapper was used to find the targets matched with the pharmacodynamic groups of sanguinarine. The gene information about PDE4D in ovarian cancer research was collected in Oncomine, and the expression level was analyzed. The A2780 and SKOV3 ovarian carcinoma cells were divided into control group and sanguinarine group. CCK8 and RT-qPCR were used to detect the effects of sanguinarine on the proliferation and PDE4D mRNA expression of A2780 and SKOV3 cells. Results PDE4D was the target matched with the pharmacodynamic group of sanguinarine. There were 6 studies referred to PDE4D in ovarian cancer and normal sample collected in the Oncomine. The expression of PDE4D in tumor tissues was significantly higher than that in normal (P < 0.05). The cell experiments showed that compared with the control group, the proliferation of A2780 and SKOV3 cells was significantly inhibited and the expression of PDE4D mRNA was decreased in the sanguinarine group, with statistical significance (P < 0.05). Conclusions PDE4D gene may play a role in the occurrence and development of ovarian cancer, and sanguinarine can inhibit the growth of ovarian cancer cells, which may be related to the down-regulation of PDE4D expression. -
Key words:
- Ovarian cancer /
- PDE4D /
- Oncomine /
- Sanguinarine
-

图 1 Oncomine数据库中PDE4D在卵巢癌中的表达
Figure 1. Expression of PDE4D in ovarian cancer in Oncomine
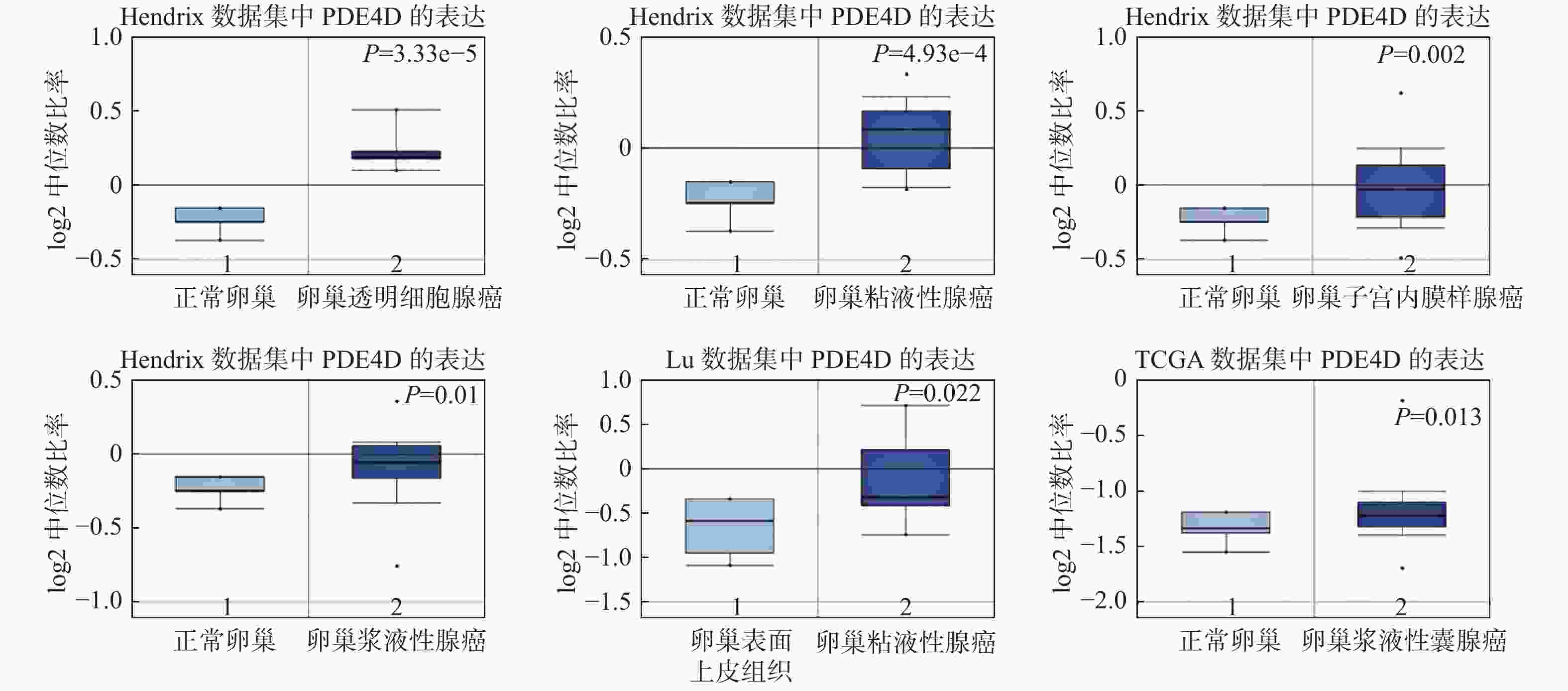
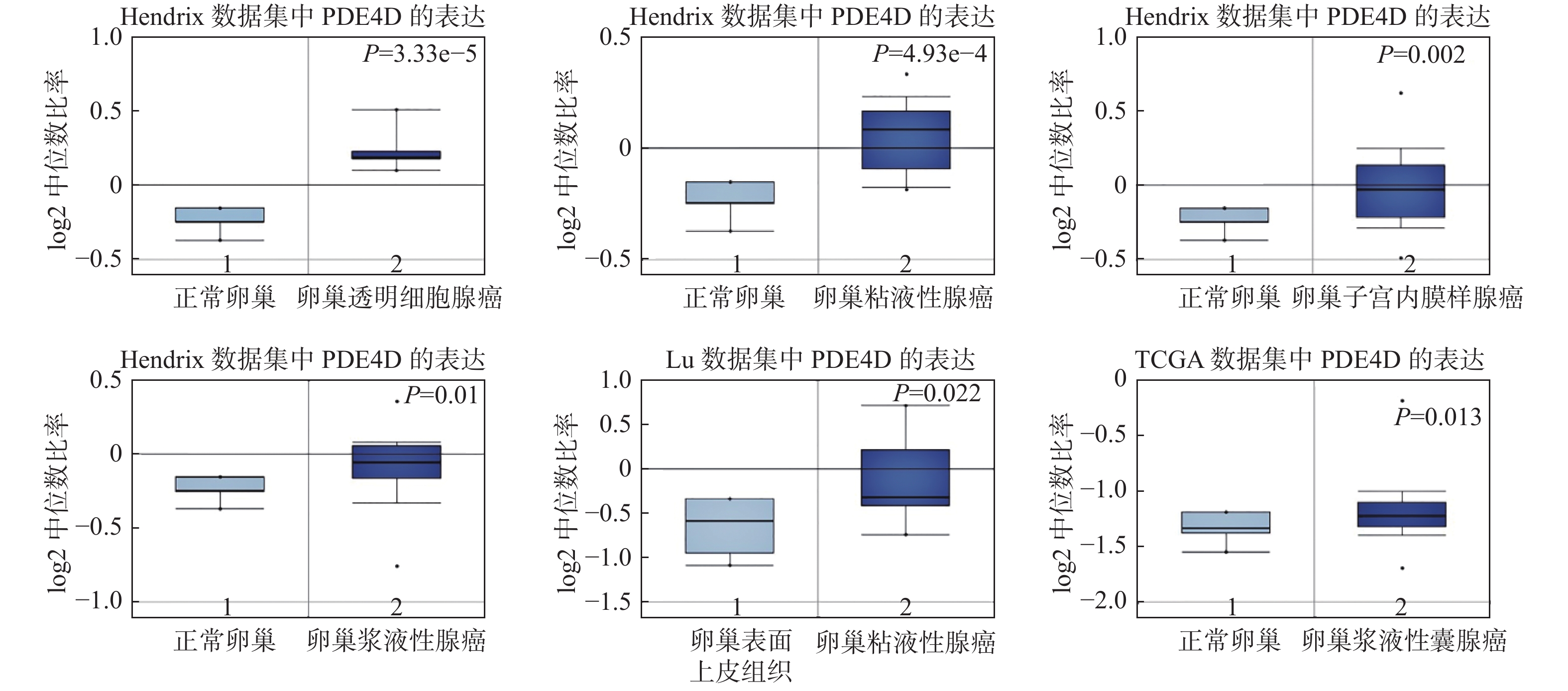
图 2 Oncomine数据库中PDE4D在不同卵巢癌芯片中的表达
Figure 2. Expression of PDE4D in different ovarian cancer studies in Oncomine
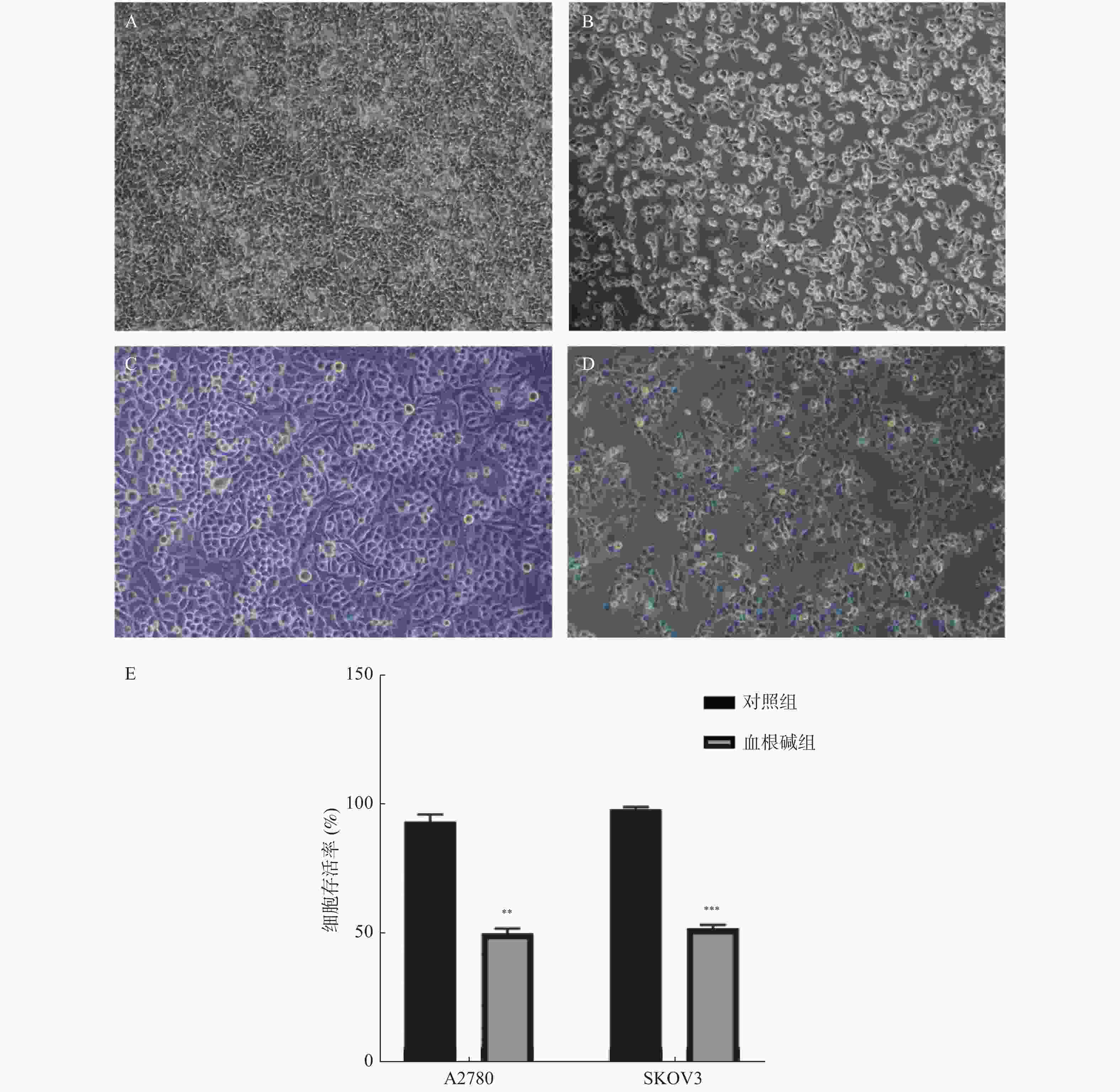
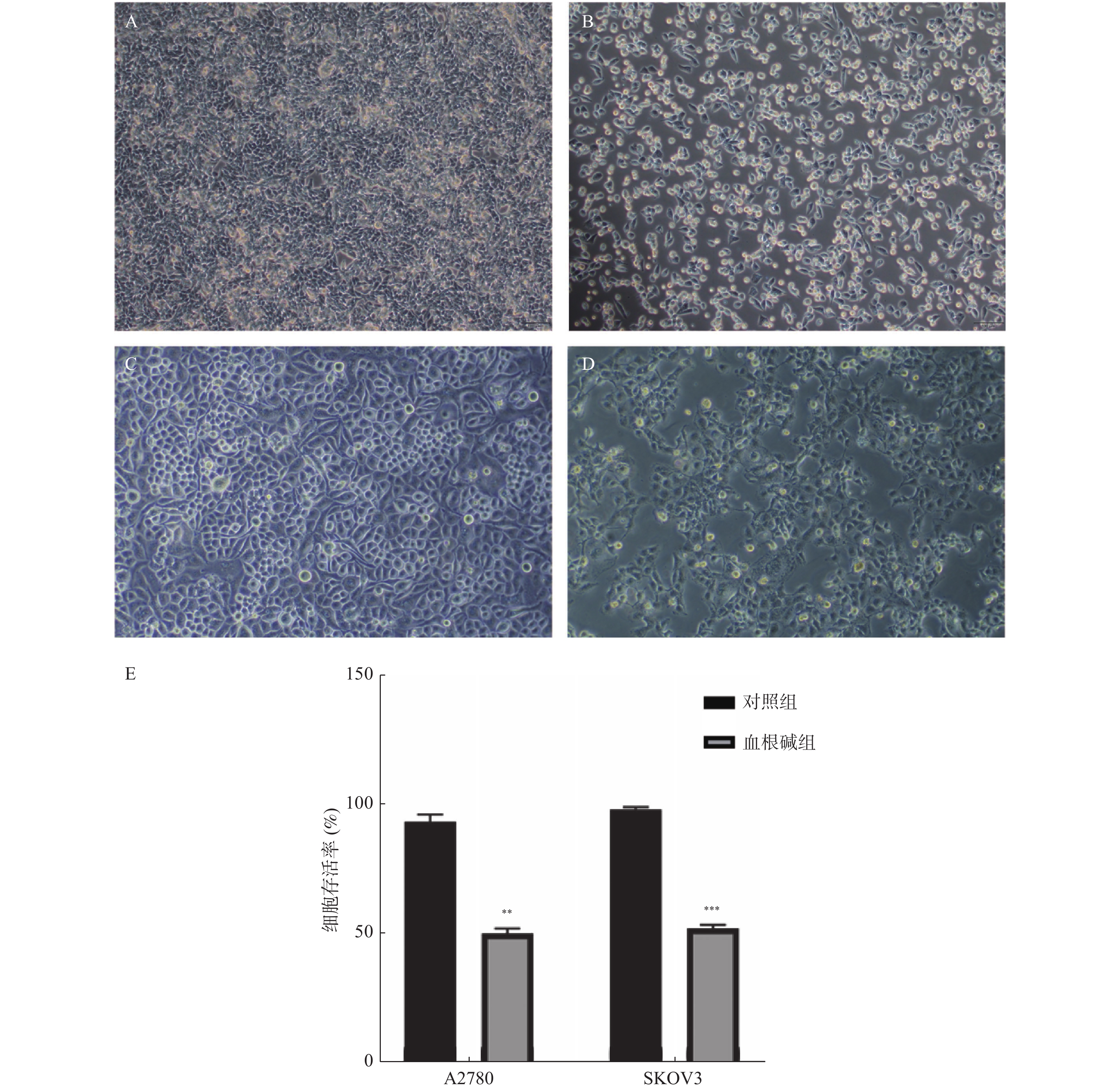
图 3 血根碱对卵巢癌A2780、SKOV3细胞增殖的影响
A:对照组A2780细胞(×100);B:血根碱组A2780细胞(×100);C:对照组SKOV3细胞(×100);D:血根碱组SKOV3细胞(×100);E:CCK8法检测的细胞存活率。与对照组比较,**P < 0.01;***P < 0.001。
Figure 3. Effect of Sanguinarine on proliferation of A2780 and SKOV3 cells
-
[1] Zhang Q,Madden N E,Wong A S T,et al. The role of endocrine G protein-coupled receptors in ovarian cancer progression[J]. Frontiers in Endocrinology,2017,8:66. [2] Hsien Lai S,Zervoudakis G,Chou J,et al. PDE4 subtypes in cancer[J]. Oncogene,2020,39(19):3791-3802. doi: 10.1038/s41388-020-1258-8 [3] Fu C,Guan G,Wang H. The anticancer effect of sanguinarine:A review[J]. Current Pharmaceutical Design,2018,24(24):2760-2764. doi: 10.2174/1381612824666180829100601 [4] Liu X, Ouyang S, Yu B, et al. Pharm Mapper server: a web server for potential drug target identification using pharmacophore mapping approach[J]. Nucleic Acids Research, 2010, 38 (Web Server issue): W609-614. [5] Rhodes D R,Yu J,Shanker K,et al. ONCOMINE:a cancer microarray database and integrated data-mining platform[J]. Neoplasia (New York,NY),2004,6(1):1-6. doi: 10.1016/S1476-5586(04)80047-2 [6] Yu H,Touna A,Yin X,et. al. Identification of differentially expressed genes and biological pathways in sanguinarine-treated ovarian cancer by integrated bioinformatics analysis[J]. Pharmacognosy Magazine,2021,17(73):106-113. doi: 10.4103/pm.pm_111_20 [7] 李京辉,朱明,曲海,等. miR-490-3p调控SW1990胰腺癌细胞上皮间充质转化[J]. 昆明医科大学学报,2021,42(3):10-17. doi: 10.12259/j.issn.2095-610X.S20210305 [8] Lin D C,Xu L,Ding L W,et al. Genomic and functional characterizations of phosphodiesterase subtype 4D in human cancers[J]. Proceedings of the National Academy of Sciences of the United States of America,2013,110(15):6109-6114. doi: 10.1073/pnas.1218206110 [9] Pullamsetti S S,Banat G A,Schmall A,et al. Phosphodiesterase-4 promotes proliferation and angiogenesis of lung cancer by crosstalk with HIF[J]. Oncogene,2013,32(9):1121-1134. doi: 10.1038/onc.2012.136 [10] Rahrmann E P,Collier L S,Knutson T P,et al. Identification of PDE4D as a proliferation promoting factor in prostate cancer using a Sleeping Beauty transposon-based somatic mutagenesis screen[J]. Cancer Research,2009,69(10):4388-4397. doi: 10.1158/0008-5472.CAN-08-3901 [11] Liu F,Ma J,Wang K,et al. High expression of PDE4D correlates with poor prognosis and clinical progression in pancreaticductal adenocarcinoma[J]. Journal of Cancer,2019,10(25):6252-6260. doi: 10.7150/jca.35443 [12] Cao B,Wang K,Liao J M,et al. Inactivation of oncogenic cAMP-specific phosphodiesterase 4D by miR-139-5p in response to p53 activation[J]. eLife,2016,5:e15978. doi: 10.7554/eLife.15978 [13] Muntean D M,Sturza A,Pavel I Z,et al. Modulation of cancer metabolism by phytochemicals-a brief overview[J]. Anti-cancer Agents in Medicinal Chemistry,2018,18(5):684-692. doi: 10.2174/1871520617666171114102218 [14] 梁宏. 基于生物信息学挖掘卵巢癌顺铂耐药机制及潜在治疗药物 [D]. 昆明: 昆明医科大学硕士论文, 2016. [15] Yu Y,Luo Y,Fang Z,et al. Mechanism of sanguinarine in inhibiting macrophages to promote metastasis and proliferation of lung cancer via modulating the exosomes in A549 cells[J]. OncoTargets and Therapy,2020,13:8989-9003. doi: 10.2147/OTT.S261054 [16] Rahman A,Pallichankandy S,Thayyullathil F,et al. Critical role of H(2)O(2) in mediating sanguinarine-induced apoptosis in prostate cancer cells via facilitating ceramide generation,ERK1/2 phosphorylation,and Par-4 cleavage[J]. Free Radical Biology & Medicine,2019,134:527-544. [17] Khan A Q,Mohamed E A N,Hakeem I,et al. Sanguinarine induces apoptosis in papillary thyroid cancer cells via generation of reactive oxygen species[J]. Molecules (Basel,Switzerland),2020,25(5):1229. doi: 10.3390/molecules25051229 [18] Fan H N,Chen W,Peng S Q,et al. Sanguinarine inhibits the tumorigenesis of gastric cancer by regulating the TOX/DNA-PKcs/ KU70/80 pathway[J]. Pathology,Research and Practice,2019,215(11):152677. [19] Gong X,Chen Z,Han Q,et al. Sanguinarine triggers intrinsic apoptosis to suppress colorectal cancer growth through disassociation between STRAP and MELK[J]. BMC Cancer,2018,18(1):578. doi: 10.1186/s12885-018-4463-x [20] Ma Y,Yu W,Shrivastava A,et al. Sanguinarine inhibits pancreatic cancer stem cell characteristics by inducing oxidative stress and suppressing sonic hedgehog-Gli-Nanog pathway[J]. Carcinogenesis,2017,38(10):1047-1056. doi: 10.1093/carcin/bgx070 [21] Sarkhosh-Inanlou R,Molaparast M,Mohammadzadeh A,et. al. Sanguinarine enhances cisplatin sensitivity via glutathione depletion in cisplatin-resistant ovarian cancer (A2780) cells[J]. Chemical Biology & Drug Design,2020,95(2):215-223. [22] Zhang S,Leng T,Zhang Q,et. al. Sanguinarine inhibits epithelial ovarian cancer development via regulating long non-coding RNA CASC2-EIF4A3 axis and/or inhibiting NF-κB signaling or PI3K/AKT/mTOR pathway[J]. Biomedicine & Pharmacotherapy = Biomedecine & Pharmacotherapie,2018,102:302-308. [23] 赵洪波,王妍妍,冷天艳,等. 血根碱对紫杉醇耐药卵巢癌A2780/taxol细胞生长及化疗敏感性影响的研究[J]. 实用妇产科杂志,2018,34(4):268-272. [24] Zhang H,Kong Q,Wang J,et al. Complex roles of cAMP-PKA-CREB signaling in cancer[J]. Experimental Hematology & Oncology,2020,9(1):32. [25] Huang H,Wang Y,Kandpal M,et al. FTO-dependent N (6)-methyladenosine modifications inhibit ovarian cancer stem cell self-renewal by blocking cAMP signaling[J]. Cancer Research,2020,80(16):3200-3214. doi: 10.1158/0008-5472.CAN-19-4044 [26] Gong S,Chen Y,Meng F,et al. Roflumilast enhances cisplatin-sensitivity and reverses cisplatin-resistance of ovarian cancer cells via cAMP/PKA/CREB-FtMt signalling axis[J]. Cell Proliferation,2018,51(5):e12474. doi: 10.1111/cpr.12474 -

 下载:
下载:
 点击查看大图
点击查看大图

计量
- 文章访问数: 4150
- HTML全文浏览量: 2007
- PDF下载量: 45
- 被引次数: 0
