

Study on the Detection of MMP-2,-7,-9,and -12 Enzymatic Activity Using CEACAM1-Derived Fluorescent Peptide Substrate Site 84
-
摘要:
目的 探索CEACAM1 源性多肽合成的荧光底物Site 84 对MMP-2,-7,-9,-12 酶活性的检测,并进一步探索荧光多肽底物Site 84在区分同属于MMPs明胶酶谱中MMP-2和MMP-9的应用。 方法 采用荧光酶学法观察荧光多肽底物Site 84 对MMP-2,-7,-9,-12 酶活性的检测;进一步观察底物Site 84对明胶酶谱中MMP-2和MMP-9酶活性的敏感性和特异性;获得底物Site 84与MMP-2的酶促反应的动力学参数Km、Kcat。 结果 以Site 84 为底物,获得MMP-12、-7、-2 的酶活力曲线,但未获得MMP-9的酶活力曲线;此外,以Site 84 为底物,可特异性检测明胶酶谱中MMP-2 的酶活性,其可检测低浓度(0.6 μM)MMP-2 的酶活性,然而对高浓度(6 μM)MMP-9 未见明显酶促反应;最后,MMP-2 和底物 Site 84的酶促反应动力学参数:Km = 315 μM,Kcat/Km = 2565 / MS。结论 以CEACAM1源性底物 Site 84作为新型的荧光多肽底物,可获得MMP-12、-7、-2 的酶活力曲线,且可特异性检测MMPs明胶酶谱中MMP-2的酶活性。 Abstract:Objective To explore the the detection of MMP-2, -7, -9, and -12 enzymatic activity using the CEACAM1-derived fluorescent peptide substrate Site 84, investigating the application of substrate Site 84 to distinguishing between MMP-2 and MMP-9 in the gelatinase spectrum of MMPs. Methods The fluorescent enzymatic method was employed to observe the detection of MMP-2, -7, -9, and -12 enzymatic activity using substrate Site 84; further observations were made on the sensitivity and specificity of substrate Site 84 to enzymatic activity of MMP-2 and MMP-9 within the gelatinase spectrum; the kinetic parameters (Km and Kcat) of the enzymatic reaction between substrate Site 84 and MMP-2 were obtained. Results Using Site 84 as a substrate, enzymic kinetics curves for MMP-12, -7, -2 were obtained, but no enzymatic activity curve for MMP-9 was observed. Furthermore, Site 84 specifically detected the enzymatic activity of MMP-2 within the gelatinase spectrum, capable of detecting low concentration (0.6 μM) of MMP-2 enzymatic activity, but no obvious enzymatic reaction was observed for high concentration (6 μM) of MMP-9; the kinetics parameters for the enzymatic reaction between Site 84 and MMP-2 were Km = 315 μM, Kcat/Km = 2565 /MS.Conclusion The CEACAM1-derived substrate Site 84 serves as a novel fluorescent peptide substrate, enabling the acquisition of enzymatic activity curves for MMP-12, -7 and -2, and specifically detecting the enzymatic activity of MMP-2 within the MMP gelatinase spectrum. -
Key words:
- Site 84 /
- Fluorescent polypeptide substrate /
- Enzymatic activity /
- Gelatinase /
- CEACAM1-derived.
-

图 1 Site84的高效液相色谱(HPLC)分析图和质谱分析图
A:HPLC纯化图,波峰即为Site 84;B:图A中波峰的二次质谱图。
Figure 1. HPLC analysis and mass spectrometry analysis of Site 84
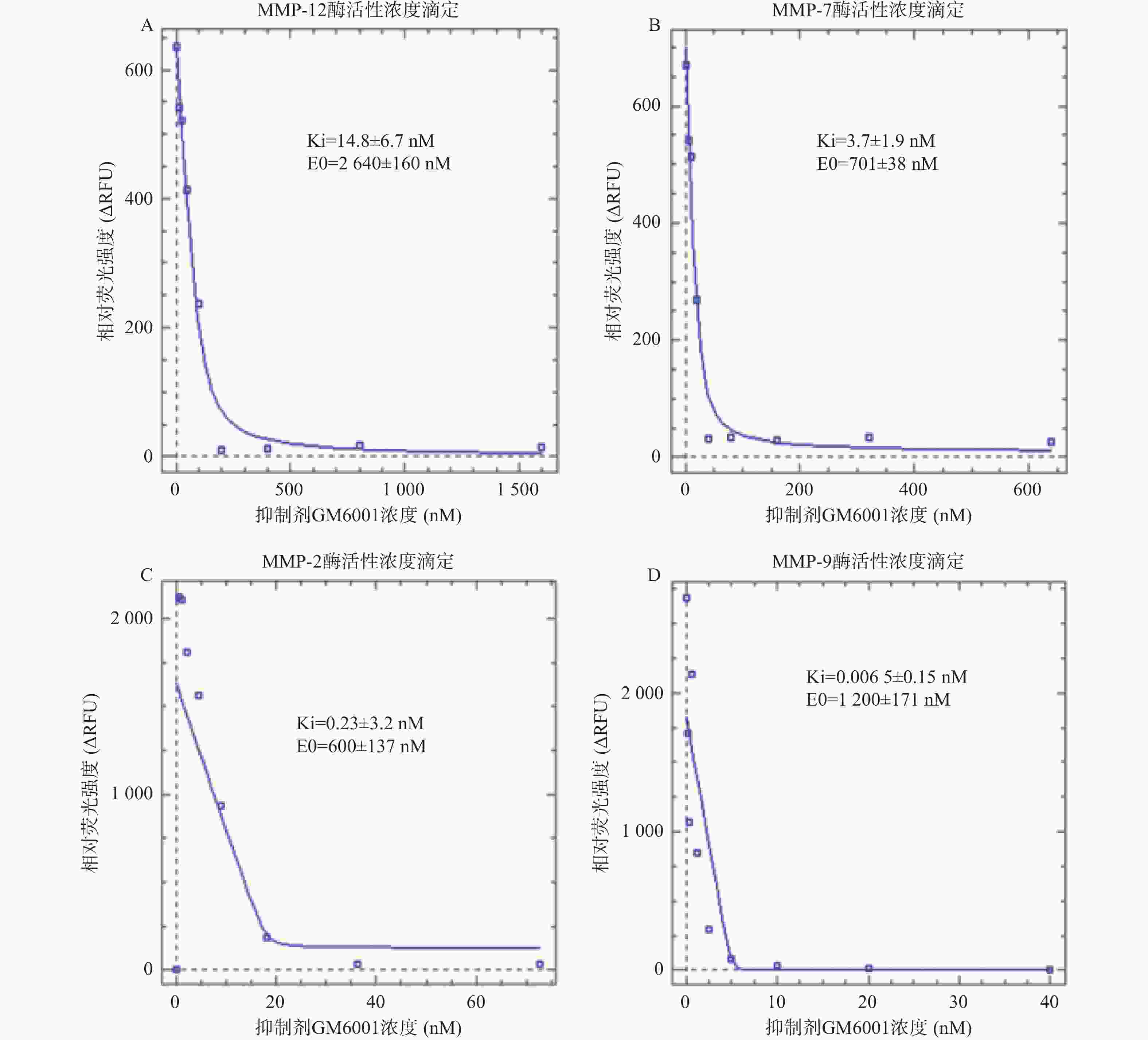
图 2 rhMMPs酶活性位点滴定
A:MMP-12的酶活性浓度;B:MMP-7的酶活性浓度;C:MMP-2的酶活性浓度;D:MMP-9的酶活性浓度。
Figure 2. Titration of enzymatic activity sites of rhMMPs
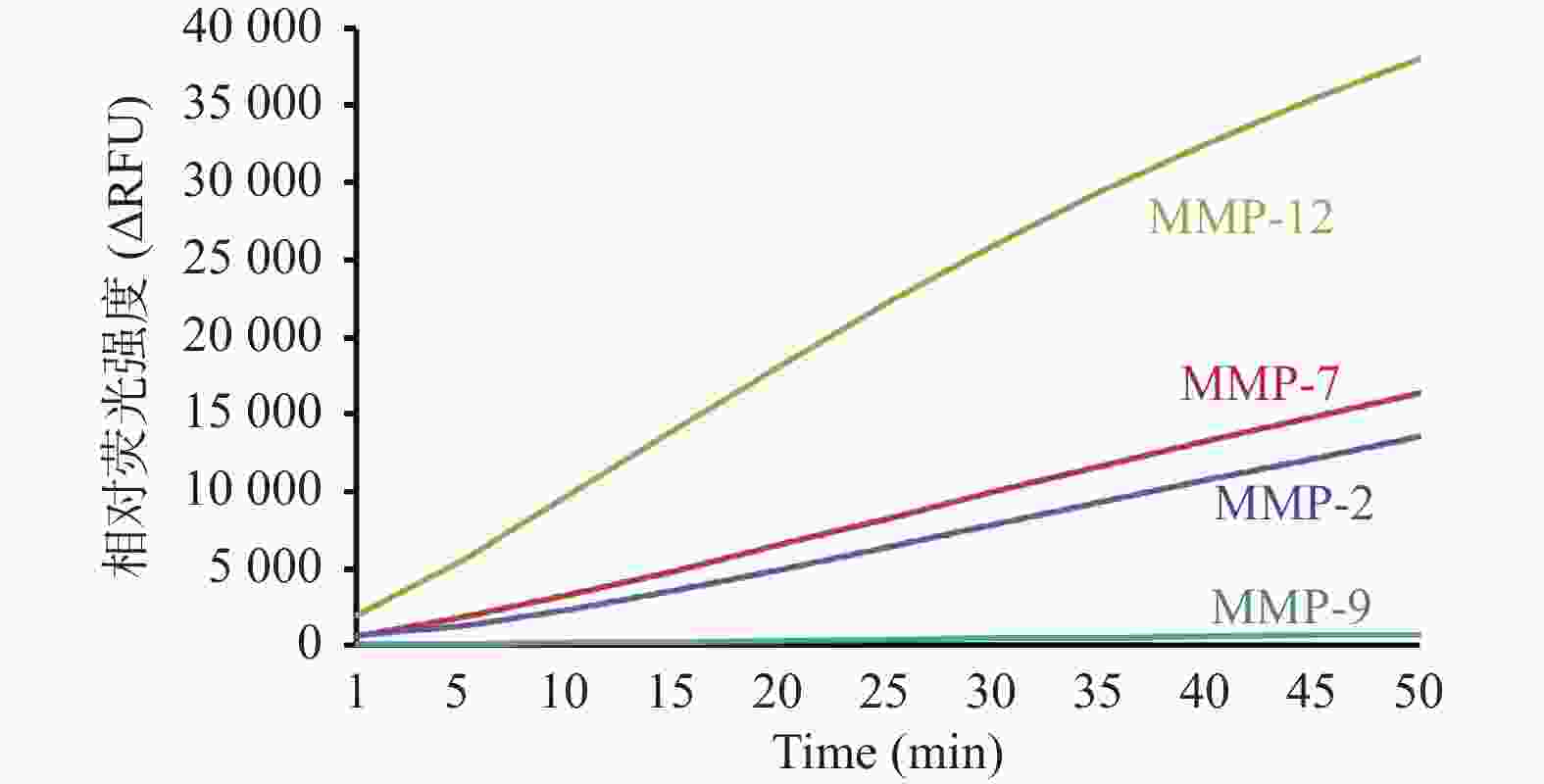
图 3 MMP-2/-7/-9/-12 与荧光多肽底物Site 84的酶促反应
Figure 3. Enzymatic reactions of MMP-2/-7/-9/-12 with fluorescent peptide substrate Site 84
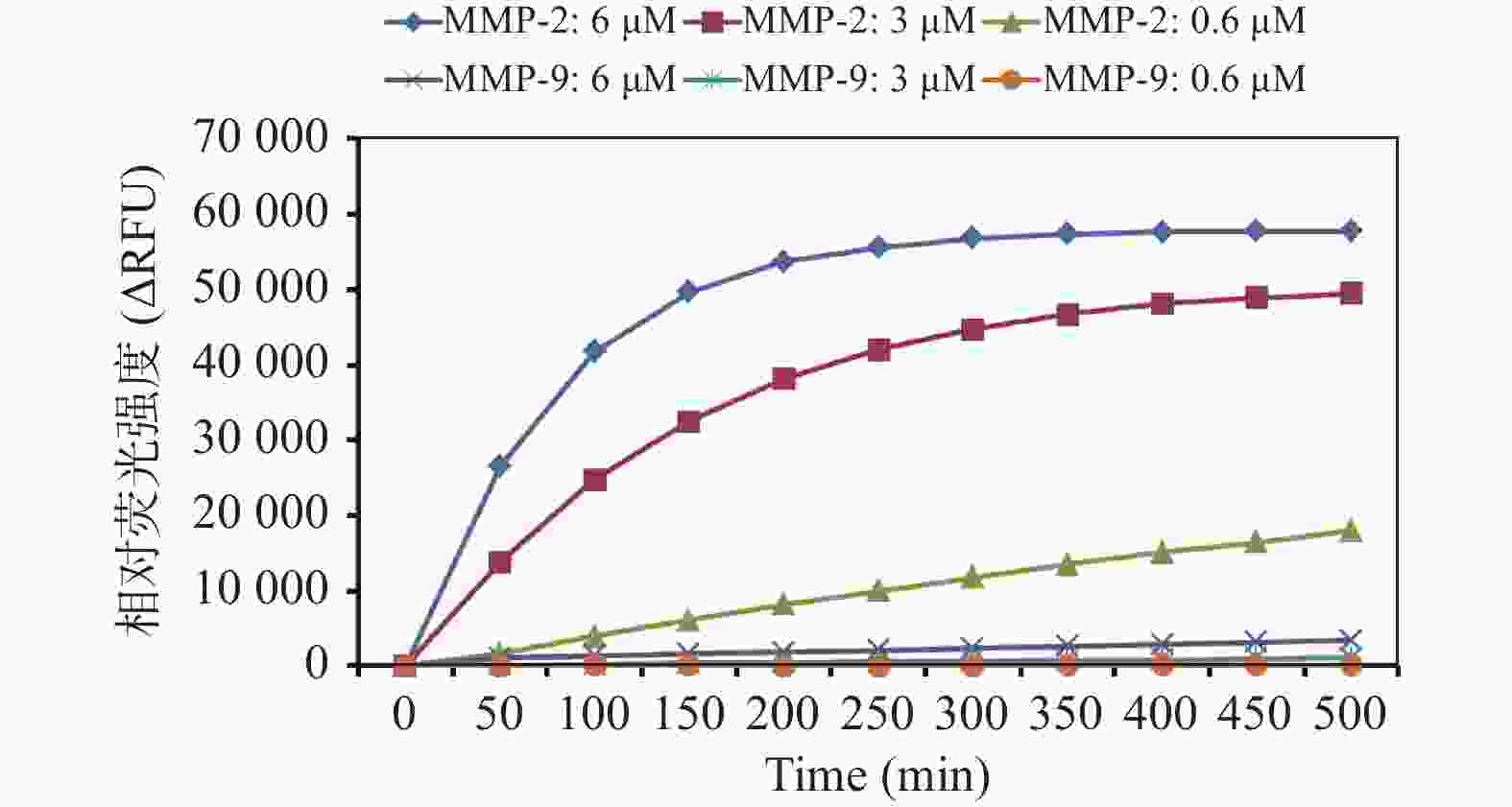
图 4 明胶酶(MMP-2 及 MMP-9)与荧光多肽底物 Site 84的酶促反应
Figure 4. Enzymatic reactions of gelatinases (MMP-2 and MMP-9) with fluorescent peptide substrate Site 84
-
[1] Wang X,Khalil R A. Matrix metalloproteinases,vascular remodeling,and vascular disease[J]. Advances in Pharmacology, 2018,81:241-330. [2] Kampoli A M,Tousoulis D,Papageorgiou N,et al. Matrix metalloproteinases in acute coronary syndromes: Current perspectives[J]. Current Topics in Medicinal Chemistry,2012,12(10):1192-1205. doi: 10.2174/1568026611208011192 [3] Wang J,Tan G J,Han L N,et al. Novel biomarkers for cardiovascular risk prediction[J]. Journal of Geriatric Cardiology: JGC,2017,14(2):135-150. [4] Dabek J,Kulach A,Gasior Z. The role of matrix metalloproteinases in acute coronary syndromes[J]. European Journal of Internal Medicine, 2007,18(6):463-466. doi: 10.1016/j.ejim.2007.01.007 [5] Ye Y,Wan W,Wang J,et al. The CEACAM1-derived peptide QLSN impairs collagen-induced human platelet activation through glycoprotein VI[J]. Bioscience,Biotechnology,and Biochemistry, 2020,84(1):85-94. [6] Wang J,Ye Y,Wei G,et al. Matrix metalloproteinase12 facilitated platelet activation by shedding carcinoembryonic antigen related cell adhesion molecule1[J]. Biochemical and Biophysical Research Communications, 2017,486(4):1103-1109. doi: 10.1016/j.bbrc.2017.04.001 [7] Stryer L. Fluorescence energy transfer as a spectroscopic ruler[J]. Annual Review of Biochemistry, 1978,47:819-846. doi: 10.1146/annurev.bi.47.070178.004131 [8] 孟照辉,叶雨佳,谢月辉,等. 一种检测人基质金属蛋白酶-12活性的荧光多肽底物[P]. 云南省: CN201410450501.X, 2017-02-01. [9] Kuzmič P. DynaFit--a software package for enzymology[J]. Methods in Enzymology, 2009,467: 247-280. [10] Santos-Martínez M J,Medina C,Jurasz P,et al. Role of metalloproteinases in platelet function[J]. Thrombosis Research, 2008,121(4):535-542. doi: 10.1016/j.thromres.2007.06.002 [11] Gresele P,Falcinelli E,Sebastiano M,et al. Matrix metalloproteinases and platelet function[J]. Progress in Molecular Biology and Translational Science, 2017,147:133-165. [12] Seizer P,May A E. Platelets and matrix metalloproteinases[J]. Thrombosis and Haemostasis, 2013,110(5):903-909. [13] Visse R,Nagase H. Matrix metalloproteinases and tissue inhibitors of metalloproteinases: structure,function,and biochemistry[J]. Circulation Research, 2003,92(8):827-839. doi: 10.1161/01.RES.0000070112.80711.3D [14] Creemers E E,Cleutjens J P,Smits J F,et al. Matrix metalloproteinase inhibition after myocardial infarction: a new approach to prevent heart failure?[J]. Circulation Research, 2001,89(3):201-210. doi: 10.1161/hh1501.094396 [15] Lindsey M L,Escobar G P,Mukherjee R,et al. Matrix metalloproteinase-7 affects connexin-43 levels,electrical conduction,and survival after myocardial infarction[J]. Circulation, 2006, 113 (25):2919-2928. [16] Johnson J L,George S J,Newby A C,et al. Divergent effects of matrix metalloproteinases 3,7,9,and 12 on atherosclerotic plaque stability in mouse brachiocephalic arteries[J]. Proceedings of the National Academy of Sciences of the United States of America, 2005,102(43):15575-15580. [17] L. N,L. J,J. N,et al. Increased plasma concentration of matrix metalloproteinase-7 in patients with coronary artery disease[J]. Clinical Chemistry, 2006(8): 1522-1527. [18] Morgan A R,Rerkasem K,Gallagher P J,et al. Differences in matrix metalloproteinase-1 and matrix metalloproteinase-12 transcript levels among carotid atherosclerotic plaques with different histopathological characteristics[J]. Stroke, 2004,35(6):1310-1315. doi: 10.1161/01.STR.0000126822.01756.99 [19] Y. -B. Z,W. L,L Q. Y,et al. Expression changes and roles of matrix metalloproteinases in a rat model of traumatic deep vein thrombosis[J]. Chinese Journal of Traumatology - English Edition, 2010(3): 188-192. [20] Bergers G,Brekken R,McMahon G,et al. Matrix metalloproteinase-9 triggers the angiogenic switch during carcinogenesis[J]. Nature Cell Biology, 2000,2(10):737-744. doi: 10.1038/35036374 [21] O'Reilly M S,Wiederschain D,Stetler-Stevenson W G,et al. Regulation of angiostatin production by matrix metalloproteinase-2 in a model of concomitant resistance[J]. The Journal of Biological Chemistry, 1999,274(41):29568-29571. doi: 10.1074/jbc.274.41.29568 [22] Hannocks M J,Zhang X,Gerwien H,et al. The gelatinases,MMP-2 and MMP-9,as fine tuners of neuroinflammatory processes[J]. Matrix Biology: Journal of the International Society for Matrix Biology, 2019,75:102-113. doi: 10.1016/j.matbio.2017.11.007 [23] Kurzepa J,Kurzepa J,Golab P,et al. The significance of matrix metalloproteinase (MMP)-2 and MMP-9 in the ischemic stroke[J]. The International Journal of Neuroscience, 2014,124(10):707-716. doi: 10.3109/00207454.2013.872102 [24] Shevchenko A V,Golovanova O V,Konenkov V I,et al. Analysis of the gene polymorphism of matrix metalloproteinase-2 and -9 in patients with coronary heart disease[J]. Terapevticheskii Arkhiv, 2010,82(1):31-34. [25] Trivedi V,Boire A,Tchernychev B,et al. Platelet matrix metalloprotease-1 mediates thrombogenesis by activating PAR1 at a cryptic ligand site[J]. Cell, 2009,137(2):332-343. doi: 10.1016/j.cell.2009.02.018 [26] Sebastiano M,Momi S,Falcinelli E,et al. A novel mechanism regulating human platelet activation by MMP-2-mediated PAR1 biased signaling[J]. Blood, 2017,129(7):883-895. doi: 10.1182/blood-2016-06-724245 [27] Gresele P,Falcinelli E,Loffredo F,et al. Platelets release matrix metalloproteinase-2 in the coronary circulation of patients with acute coronary syndromes: possible role in sustained platelet activation[J]. European Heart Journal, 2011(3):316-325. [28] Martinez A,Salas E,Radomski A,et al. Matrix metalloproteinase-2 in platelet adhesion to fibrinogen: interactions with nitric oxide[J]. Medical Science Monitor: International Medical Journal of Experimental and Clinical research, 2001(4):646-651. [29] Sanchis L,Andrea R,Falces C,et al. Prognosis of new-onset heart failure outpatients and collagen biomarkers[J]. European Journal of Clinical Investigation, 2015,45(8):842-849. doi: 10.1111/eci.12479 [30] Sheu J R,Fong T H,Liu C M,et al. Expression of matrix metalloproteinase-9 in human platelets: regulation of platelet activation in in vitro and in vivo studies[J]. British Journal of Pharmacology, 2004,143(1):193-201. doi: 10.1038/sj.bjp.0705917 [31] Fernandez-Patron C,Martinez-Cuesta MA,Salas E,et al. Differential regulation of platelet aggregation by matrix metalloproteinases-9 and -2[J]. Thrombosis and Haemostasis, 1999,82(6):1730-1735. [32] Dini F L,Buralli S,Bajraktari G,et al. Plasma matrix metalloproteinase-9 better predicts outcome than N-terminal protype-B natriuretic peptide in patients with systolic heart failure and a high prevalence of coronary artery disease[J]. Biomedicine & Pharmacotherapy, 2010,64(5):339-342. [33] Wagner D R,Delagardelle C,Ernens I,et al. Matrix metalloproteinase-9 is a marker of heart failure after acute myocardial infarction[J]. Journal of Cardiac Failure, 2006,12(1):66-72. doi: 10.1016/j.cardfail.2005.08.002 [34] Kelly D,Khan S Q,Thompson M,et al. Plasma tissue inhibitor of metalloproteinase-1 and matrix metalloproteinase-9: novel indicators of left ventricular remodeling and prognosis after acute myocardial infarction[J]. Eur Heart J, 2008,29(17):2116-2124. doi: 10.1093/eurheartj/ehn315 -

 下载:
下载:




 点击查看大图
点击查看大图

计量
- 文章访问数: 345
- HTML全文浏览量: 219
- PDF下载量: 20
- 被引次数: 0
