

Mechanism of Methyltransferase METTL3 in Premature Ovarian Failure
-
摘要:
目的 通过POF小鼠模型,探究甲基转移酶METTL3在POF中的作用机制。 方法 选取6~8周C57BL/6雌性小鼠,将采用D-半乳糖代谢法构建的POF小鼠模型设为实验组(n = 15),以等量生理盐水同样方式干预的正常小鼠作为对照组(n = 15)。通过ELISA检测性激素水平、HE染色观察卵巢组织病理变化,验证POF小鼠模型构建成功,并在组织水平探讨METTL3与POF的相关性;同时在细胞水平构建METTL3过表达和敲低的小鼠颗粒细胞稳转录株,利用CCK-8和FITC/PI双染法检测细胞增殖与凋亡情况,以明确METTL3对颗粒细胞凋亡的调控作用及在POF发生中的影响。 结果 成功构建POF小鼠模型,经TUNEL染色发现POF组小鼠卵巢组织中凋亡的颗粒细胞明显增多,且该组小鼠卵巢组织中的METTL3呈高表达(P < 0.05)。在细胞实验中,成功构建METTL3过表达和敲低的小鼠颗粒细胞稳转录株,上调METTL3表达可促进颗粒细胞凋亡并抑制其增殖(P < 0.05)。 结论 METTL3通过促进颗粒细胞凋亡参与POF的发生。 Abstract:Objective To explore the mechanism of methyltransferase METTL3 in premature ovarian failure (POF) using a POF mouse model, providing a theoretical basis for the pathological research and clinical treatment of POF. Methods C57BL/6 female mice aged 6 to 8 weeks were selected for the study. The experimental group consisted of mice with a premature ovarian failure (POF) model established using the D-galactose metabolic method, while the control group comprised normal mice subjected to the same intervention with an equal volume of physiological saline. The levels of sex hormones were measured using ELISA, and histopathological changes in ovarian tissue were observed through HE staining to verify the successful establishment of the POF mouse model and to explore the correlation between METTL3 and POF at the tissue level. Additionally, stable transgenic mouse granulosa cell lines with METTL3 overexpression and knockdown were constructed at the cellular level. Cell proliferation and apoptosis were assessed using CCK-8 and FITC/PI dual staining methods to clarify the regulatory role of METTL3 in granulosa cell apoptosis and its impact on the occurrence of POF. Results The POF mouse model was successfully established. TUNEL staining revealed a significant increase in apoptotic granulosa cells in the ovarian tissue of the POF group (P < 0.05), which also exhibited high expression of METTL3. In cellular experiments, stable transcriptional cell lines of mouse granulosa cells with METTL3 overexpression and knockdown were successfully constructed. Upregulation of METTL3 expression promoted granulosa cell apoptosis and inhibited their proliferation (P < 0.05). Conclusion METTL3 participates in the development of POF by promoting granulosa cell apoptosis. -
Key words:
- Premature ovarian failure /
- Mouse model /
- METTL3 /
- Granulosa cell apoptosis
-

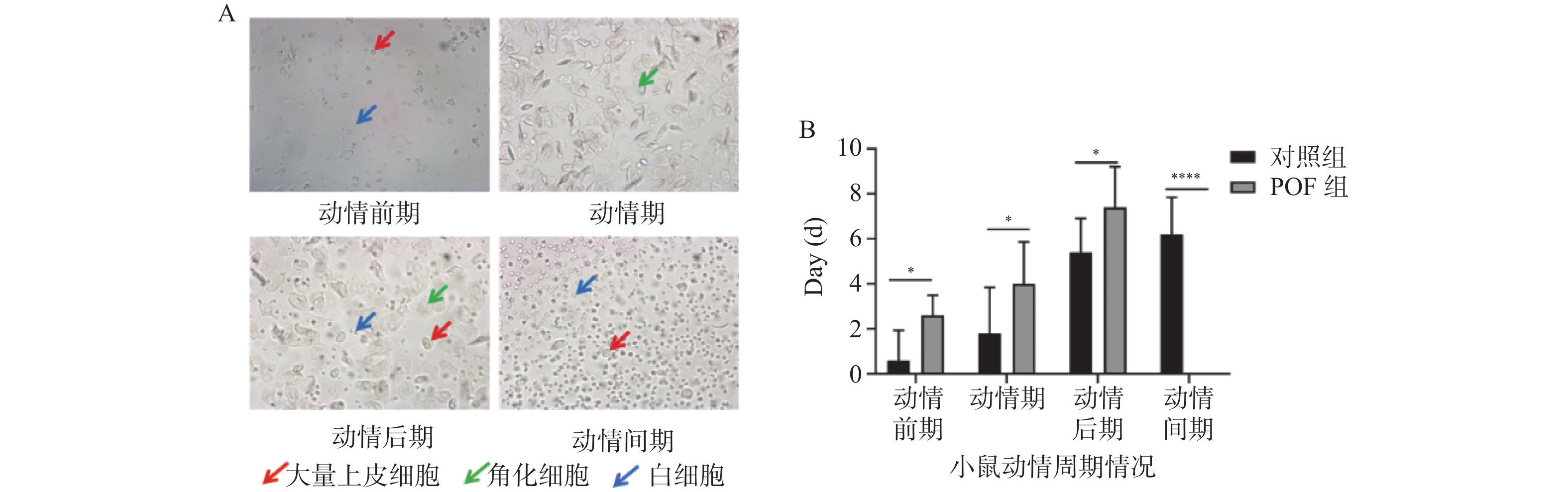
图 1 两组小鼠动情周期变化
A :两组小鼠阴道分泌物涂片周期变化; B :两组小鼠动情周期情况。*P < 0.05;****P < 0.0001。
Figure 1. Changes in the estrous cycle of mice in the two groups
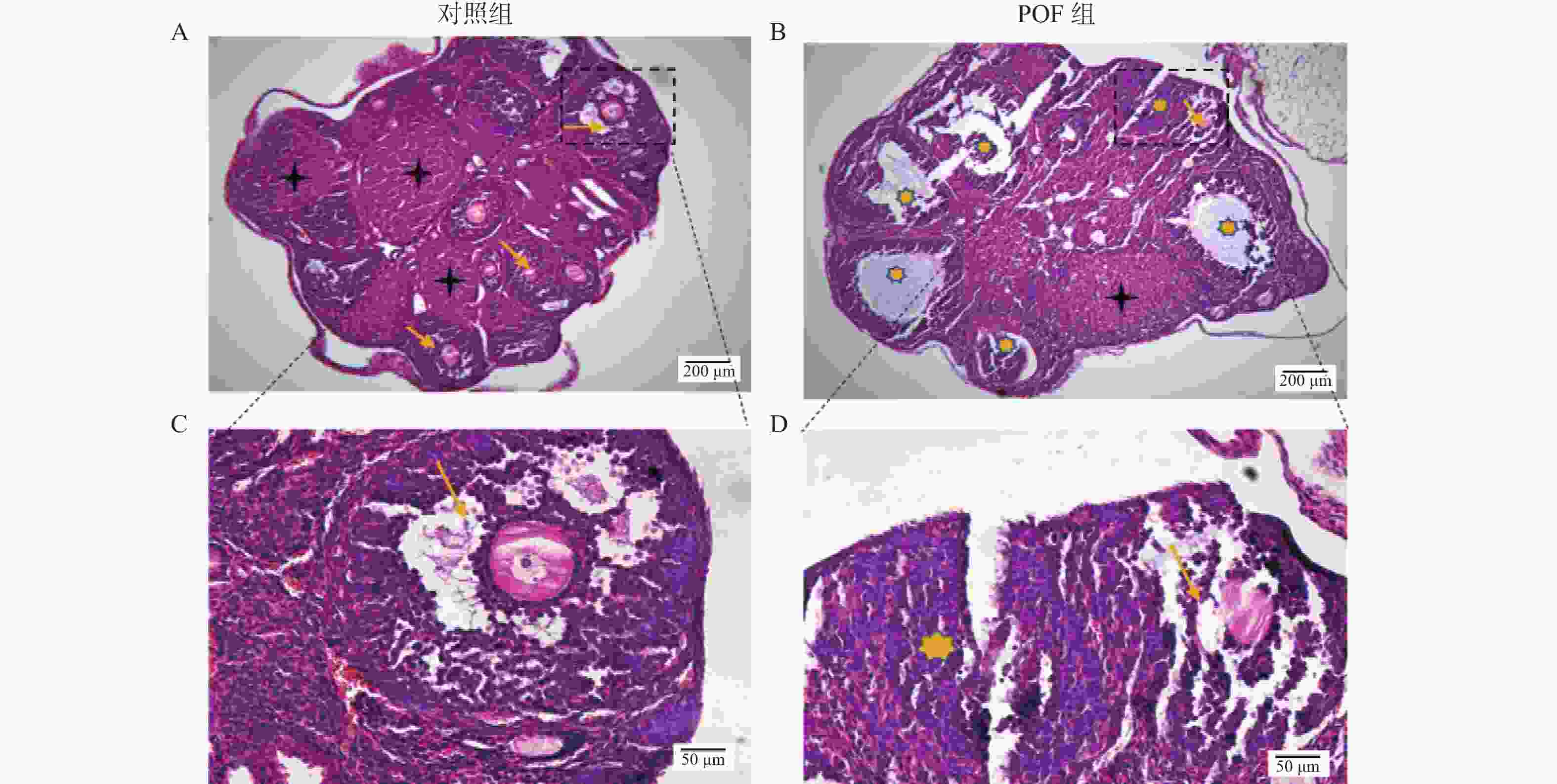
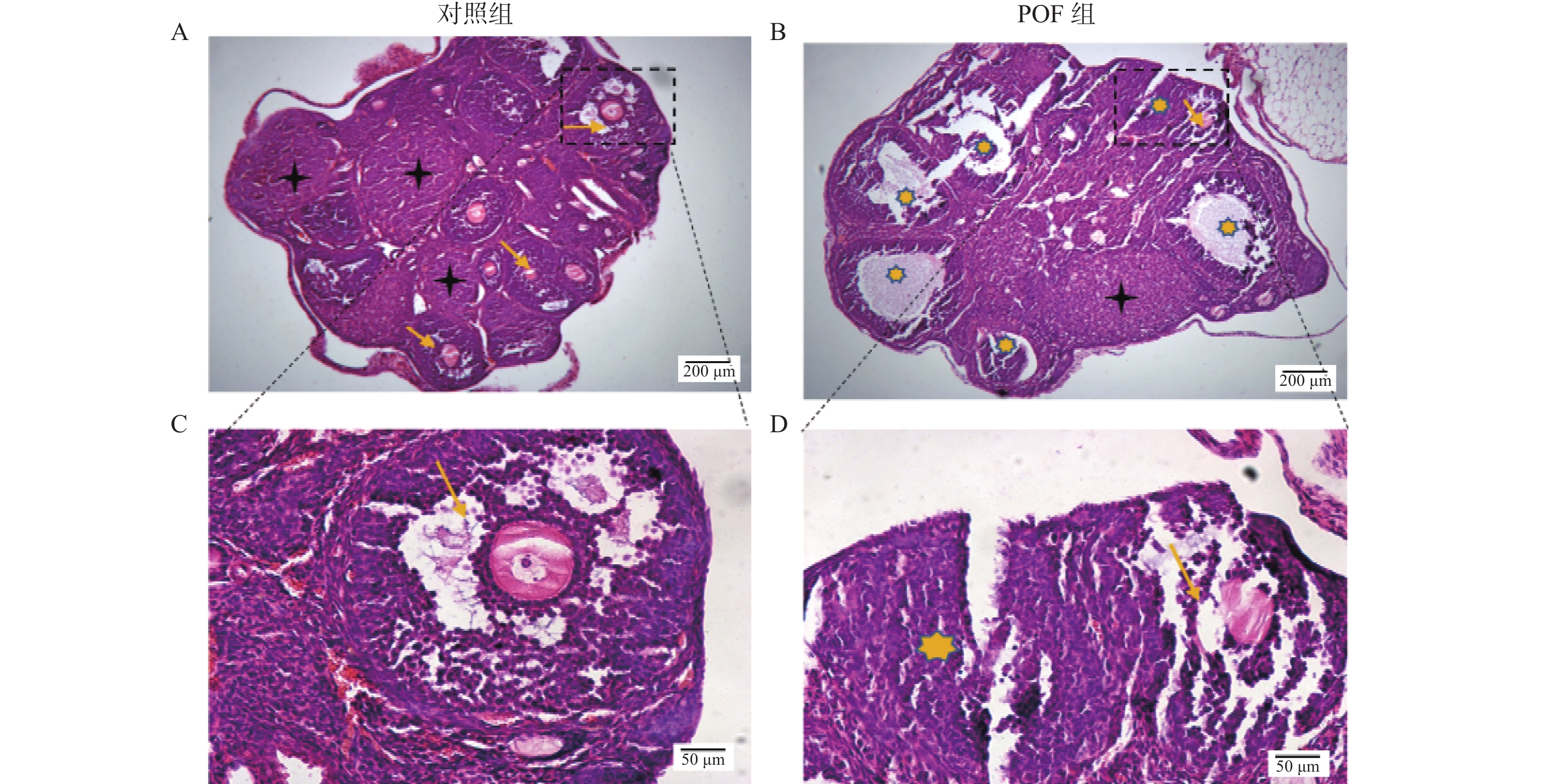
图 2 两组小鼠卵巢组织 HE染色观察小鼠卵巢组织病理变化
A:对照组小鼠卵巢组织;B:POF组小鼠卵巢组织;C:对照组小鼠卵巢组织中卵泡;D:POF组小鼠卵巢组织中卵泡。
Figure 2. Observation on the pathological changes of mouse ovarian tissues by HE staining in the two groups of mice
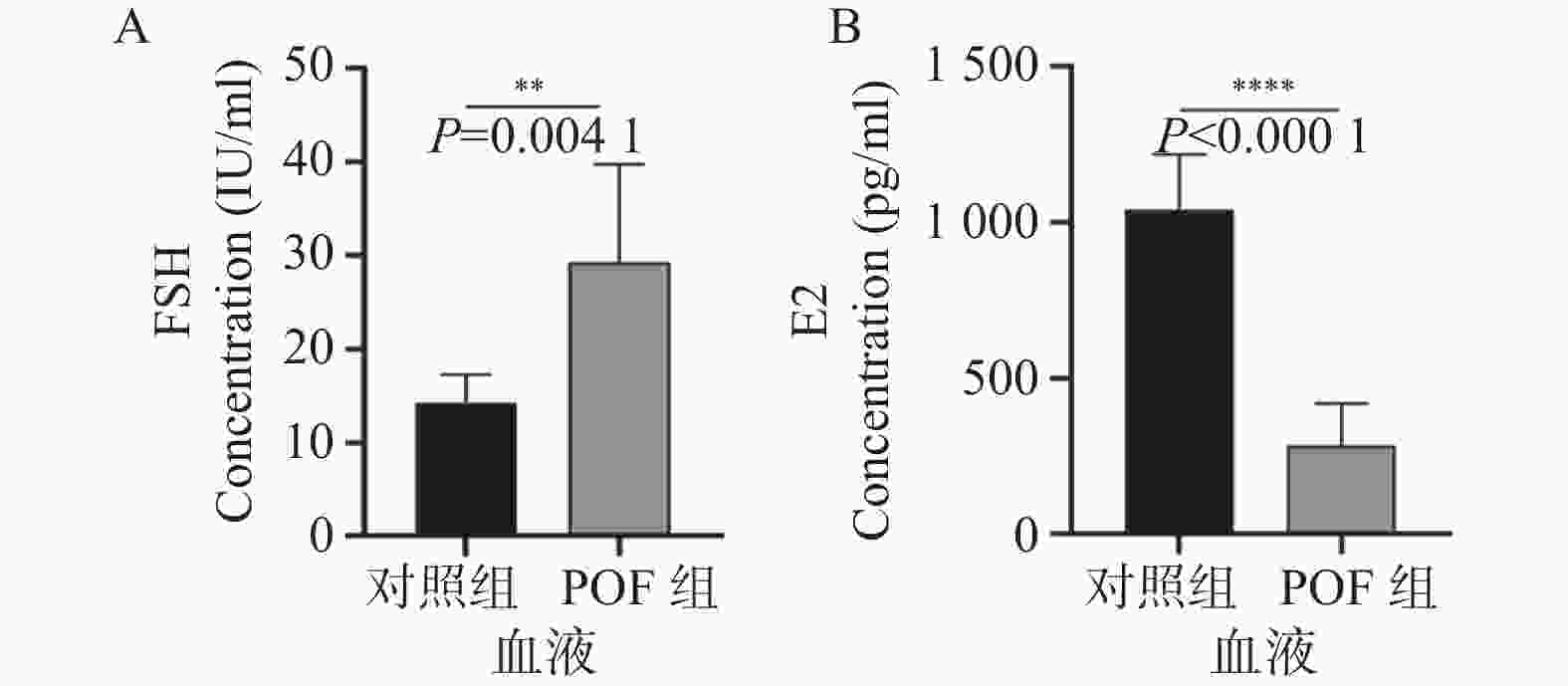
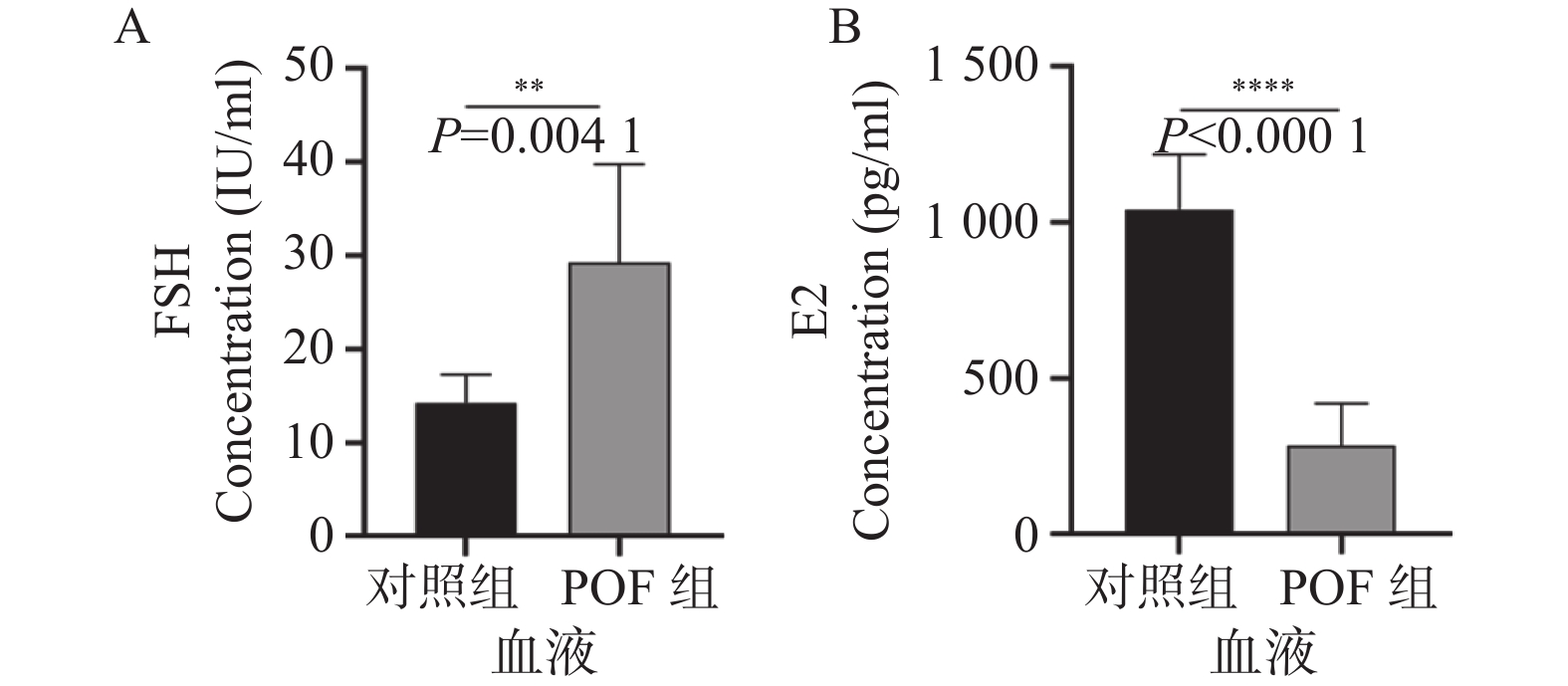
图 3 ELISA检测小鼠外周血的性激素水平结果
A:小鼠外周血的FSH激素水平变化; B:小鼠外周血的E2激素水平变化。**P < 0.01;****P < 0.0001。
Figure 3. Results of hormone level detection in mouse peripheral blood using ELISA
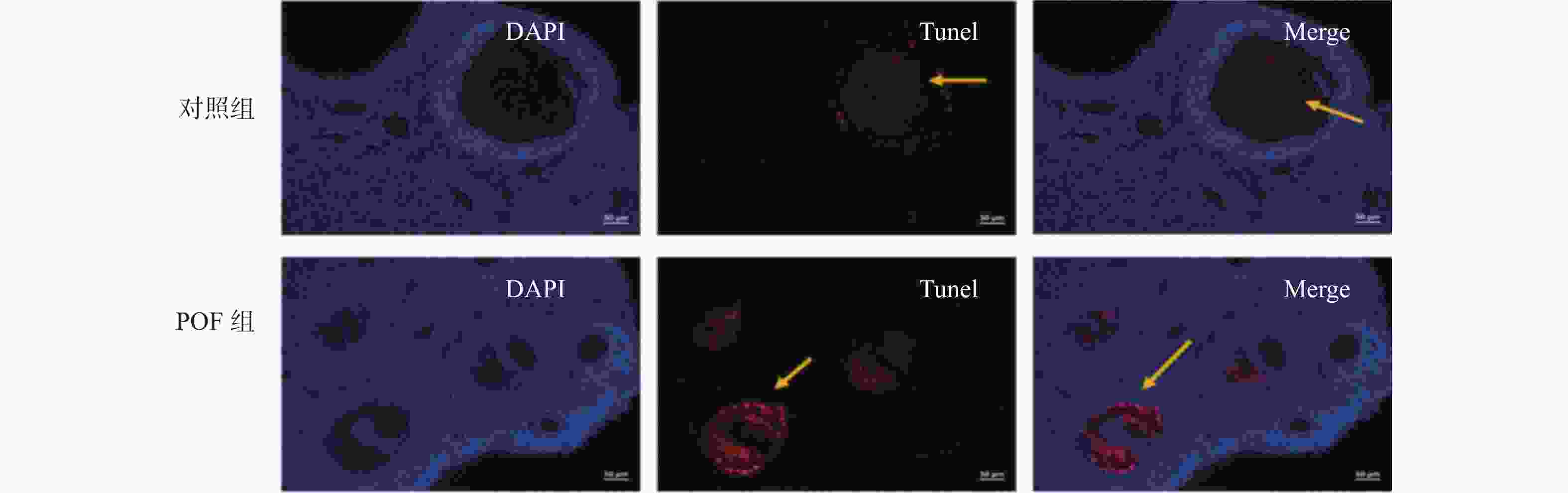
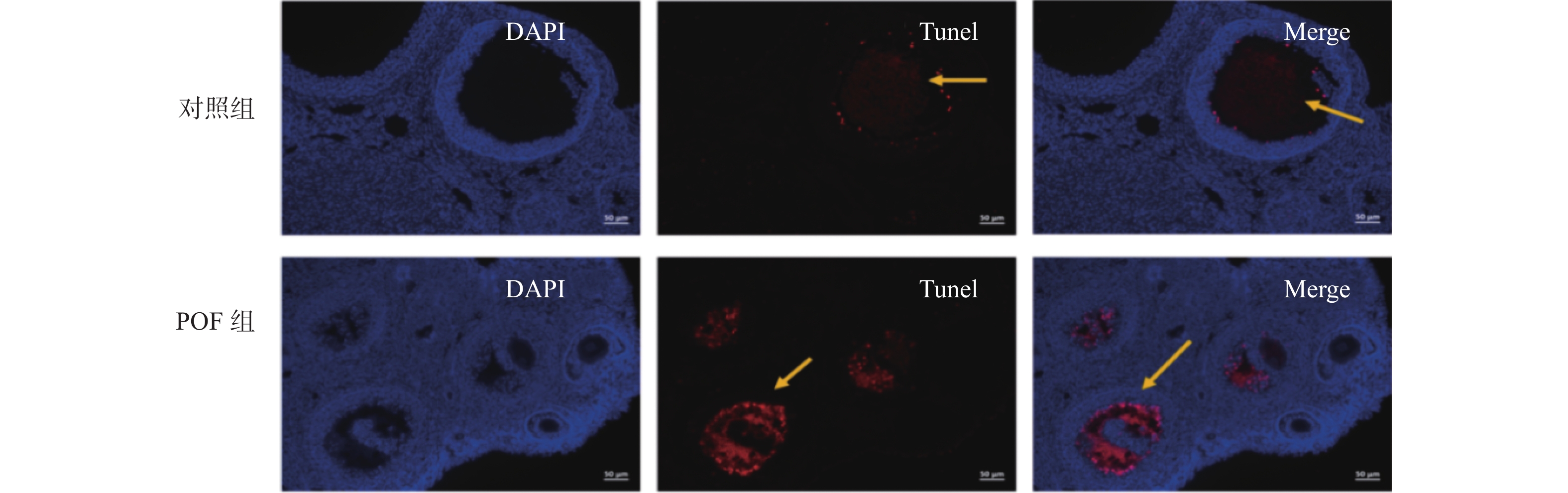
图 4 TUNEL染色检测卵巢组织细胞凋亡
Figure 4. Detection of apoptosis of ovarian tissue cells by TUNEL staining
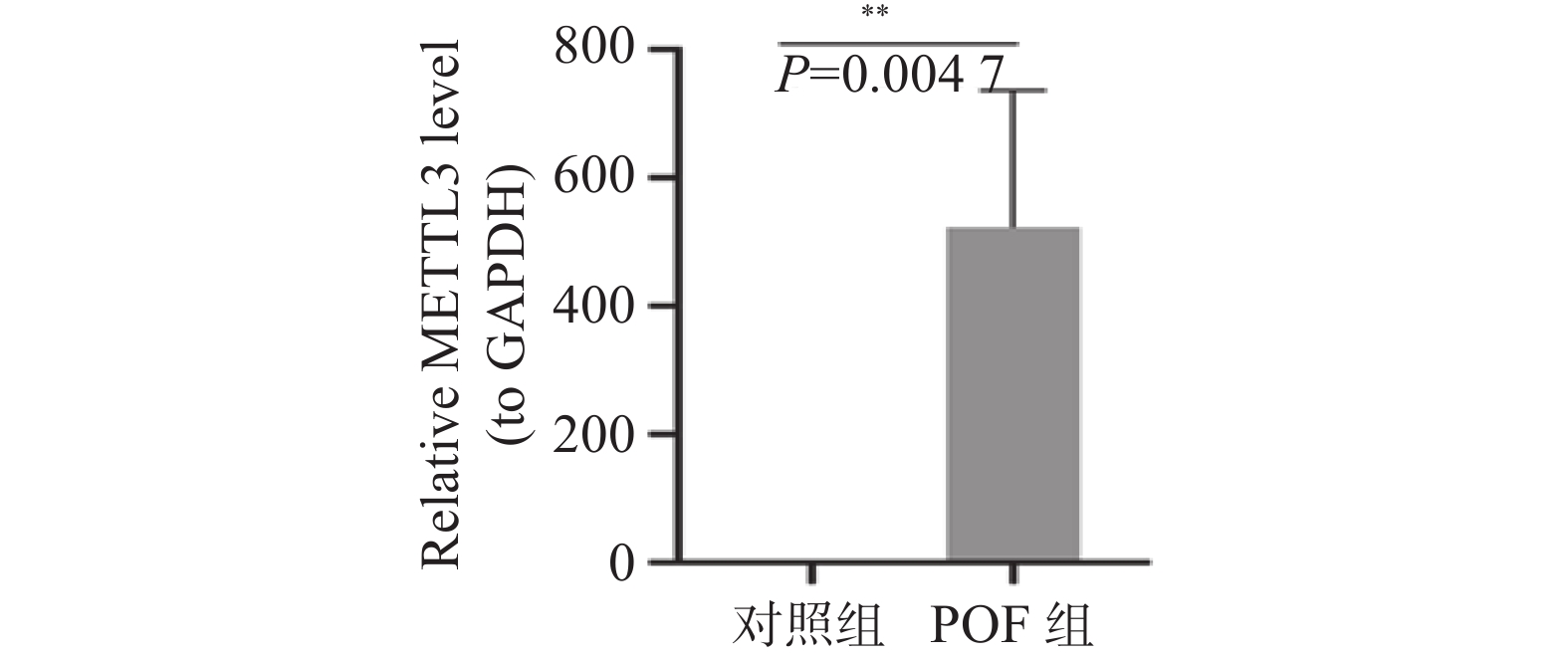
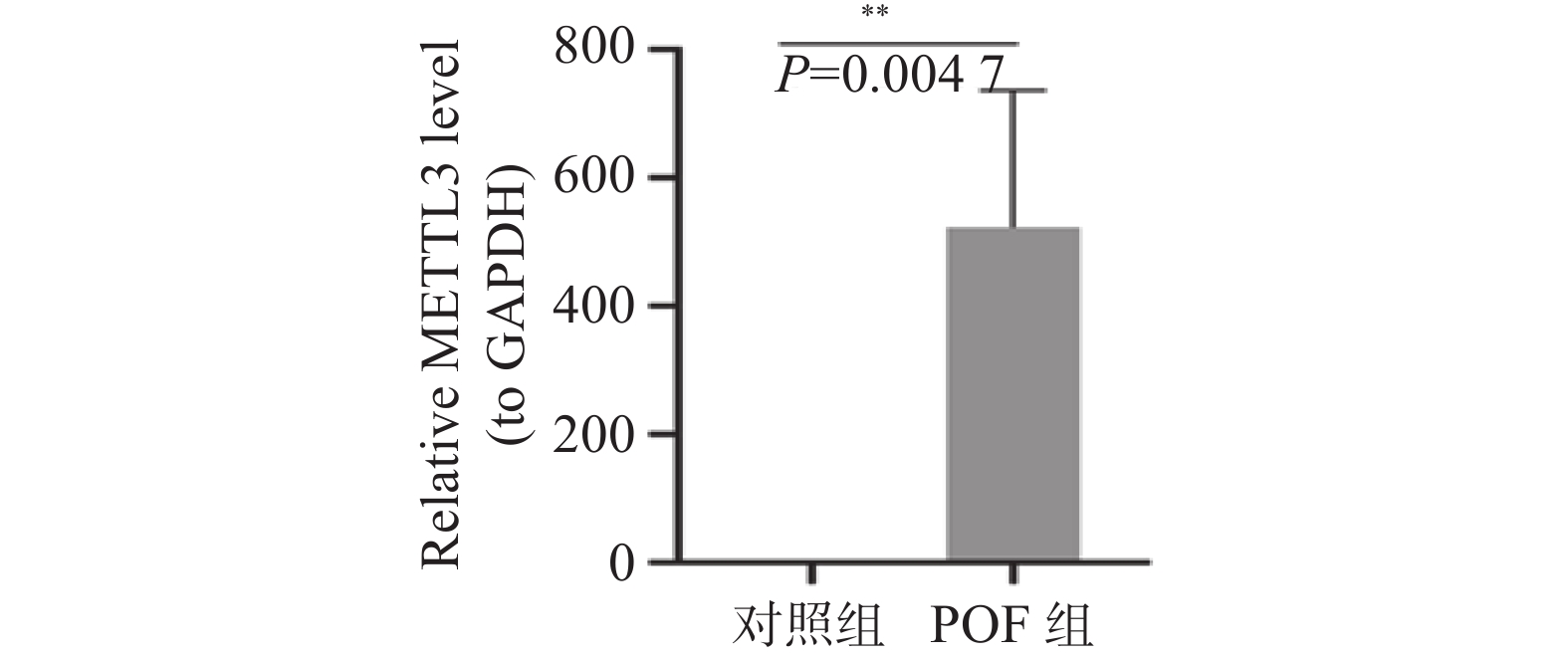
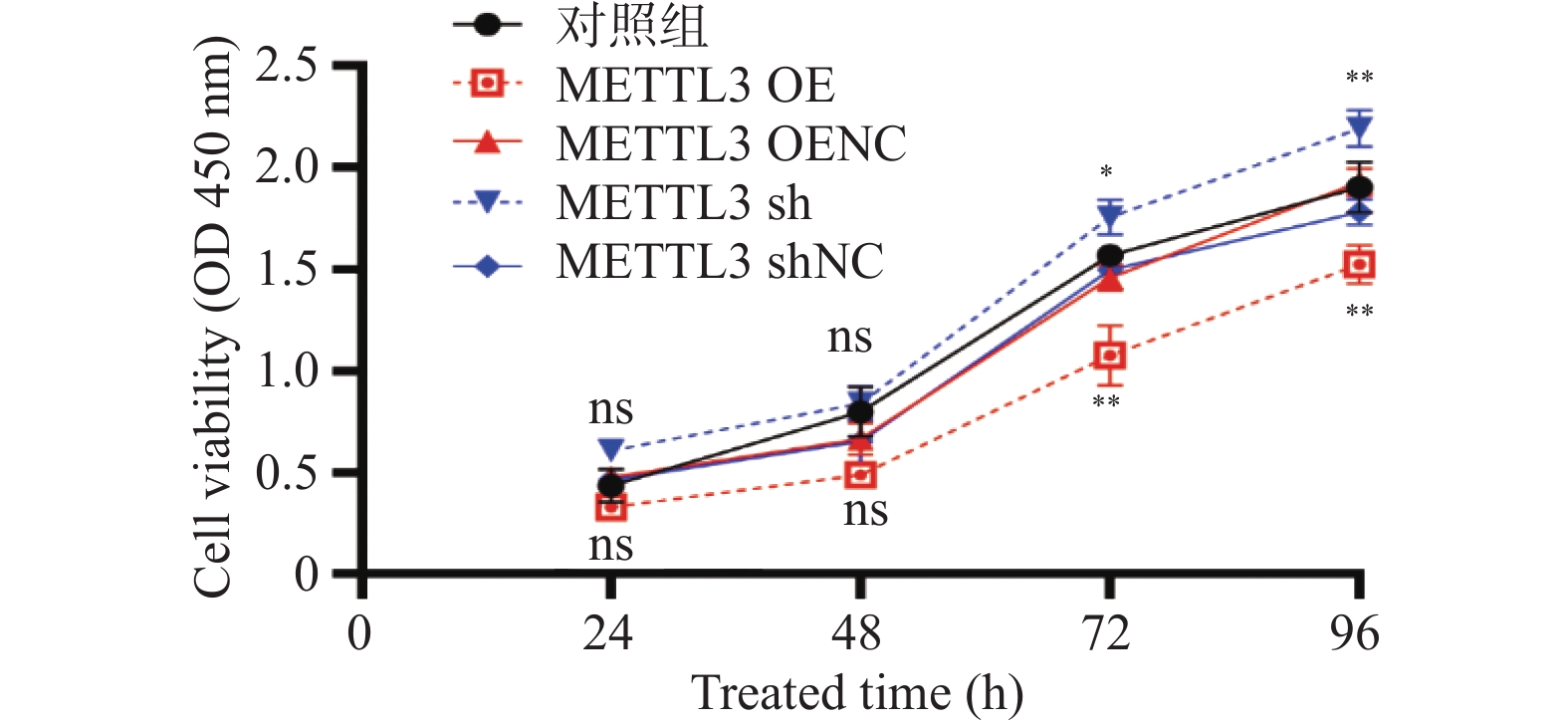
图 5 qRT-PCR检测小鼠卵巢组织METTL3的mRNA表达水平
**P < 0.01。
Figure 5. Detection of the mRNA expression level of METTL3 in mouse ovarian tissues by qRT-PCR
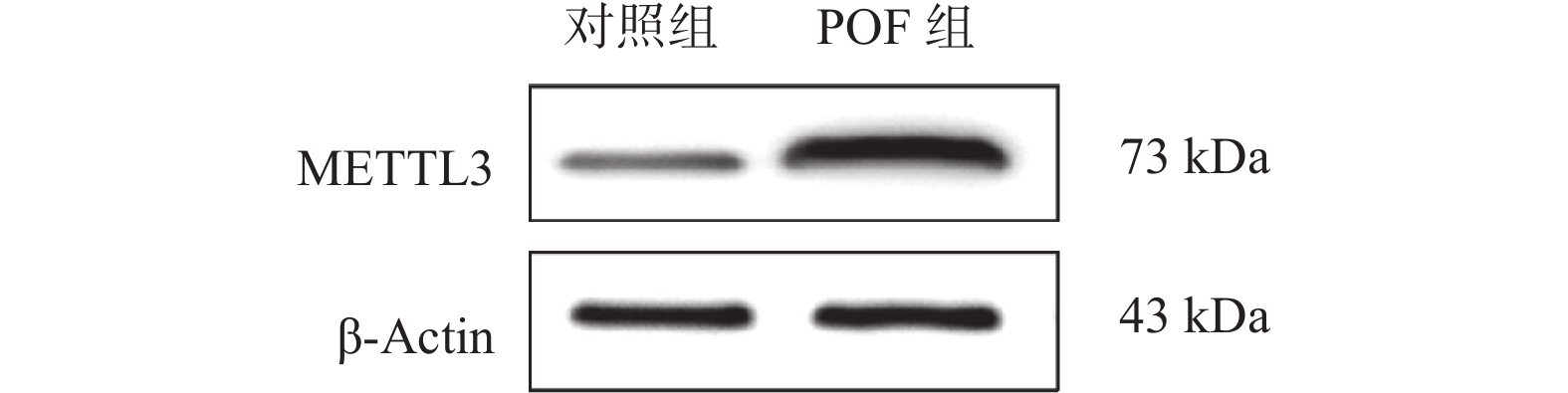
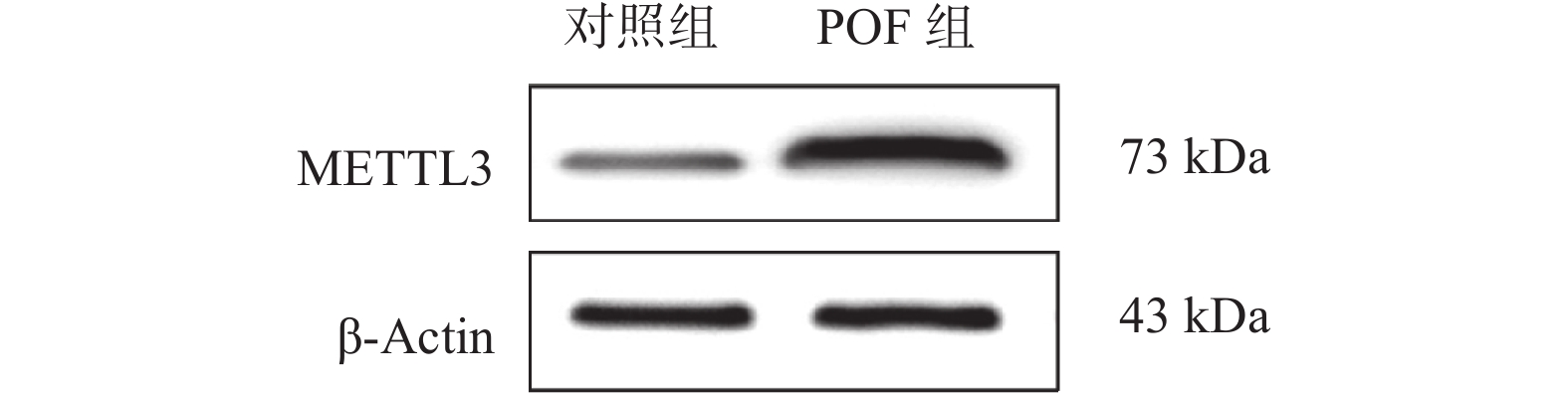
图 6 Western blots检测小鼠卵巢组织的甲基转移酶METTL3的蛋白表达水平
Figure 6. Detection of the protein expression level of the methyltransferase METTL3 in mouse ovarian tissues by Western blots
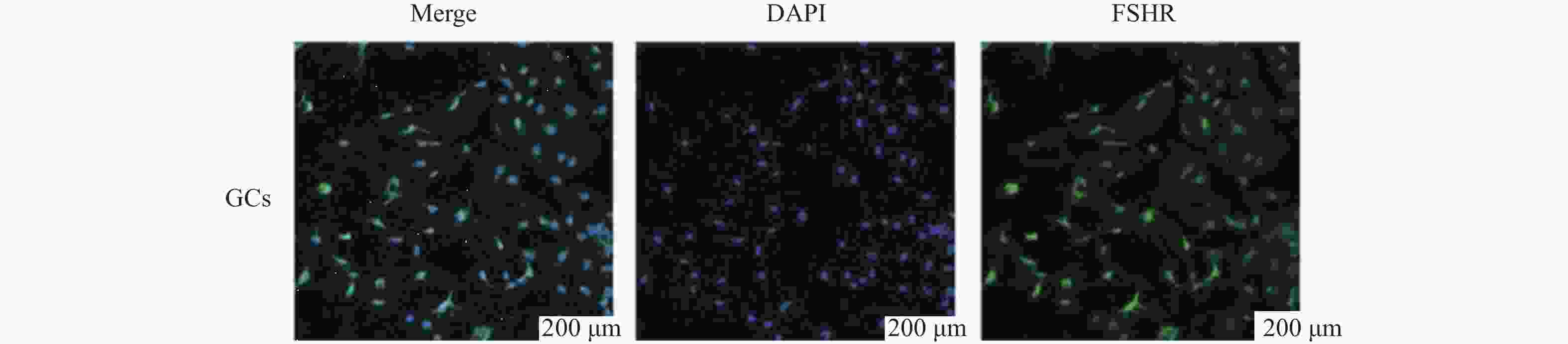
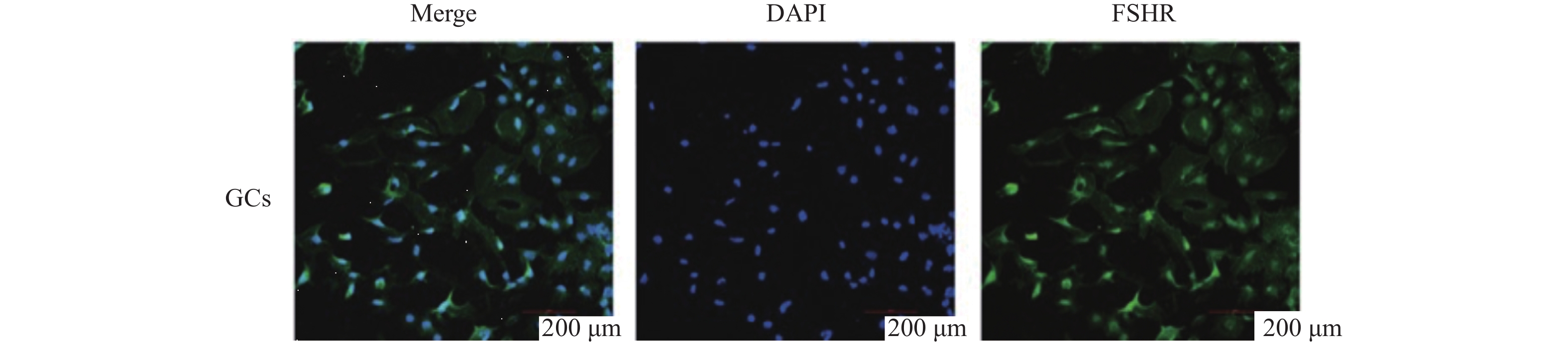
图 7 小鼠卵巢颗粒细胞染色鉴定
Figure 7. Identification of granulosa cells in mouse ovaries through staining

图 8 Western blots检测慢病毒的转染效率
Figure 8. Detection of the transfection efficiency of lentivirus by Western blots
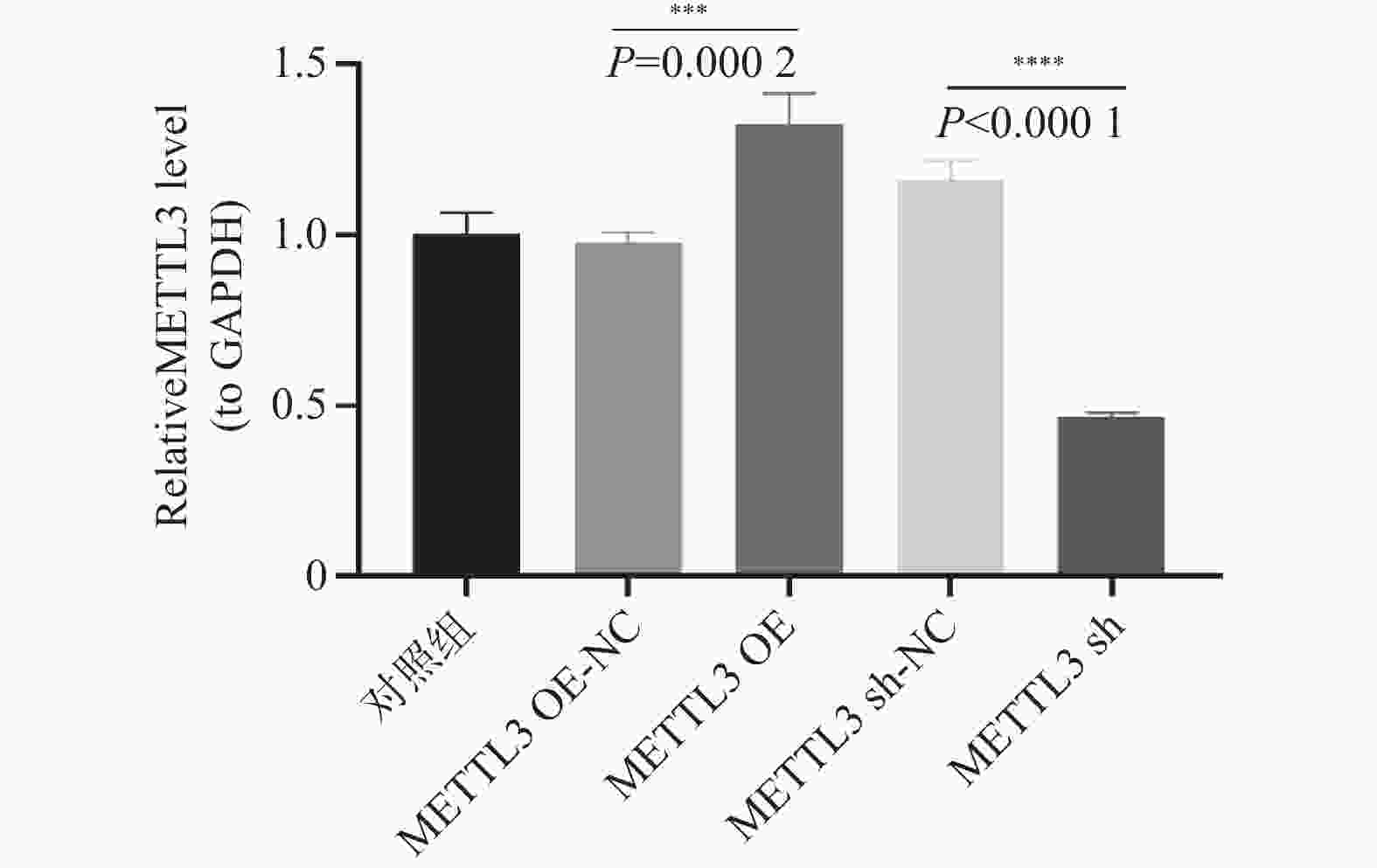
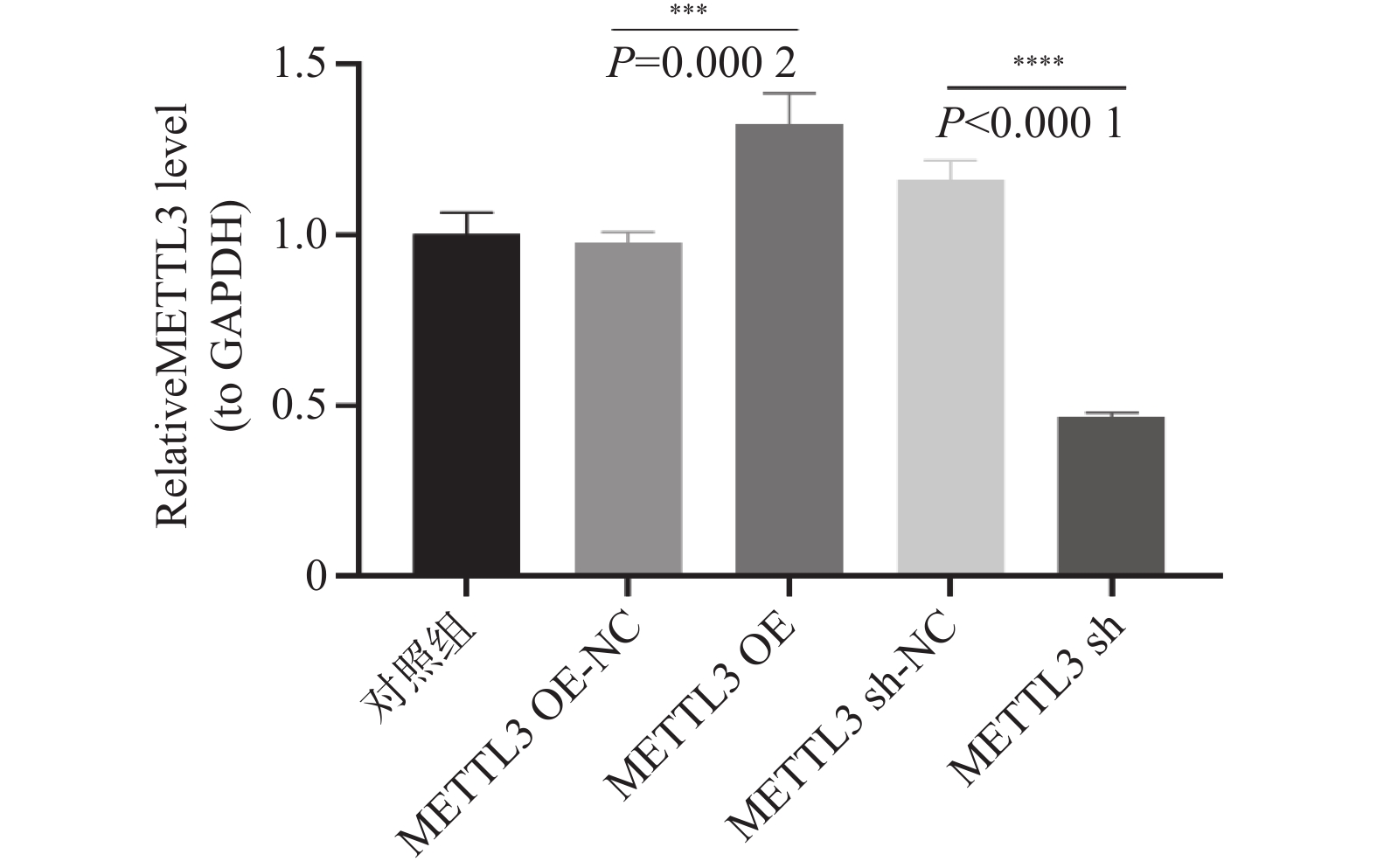
图 9 qRT-PCR检测卵巢颗粒细胞METTL3的mRNA表达水平
***P < 0.0002,****P < 0.001。
Figure 9. Detection of the mRNA expression level of METTL3 in ovarian granulosa cells by qRT-PCR
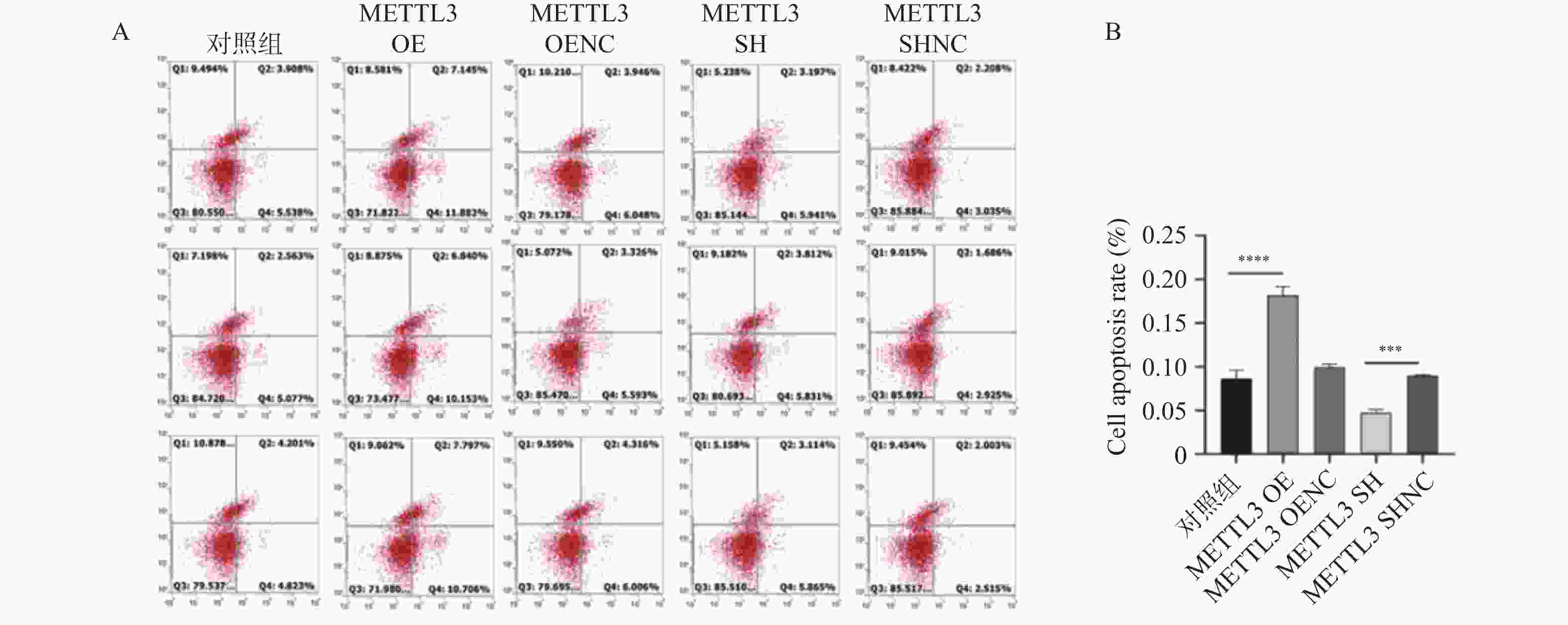
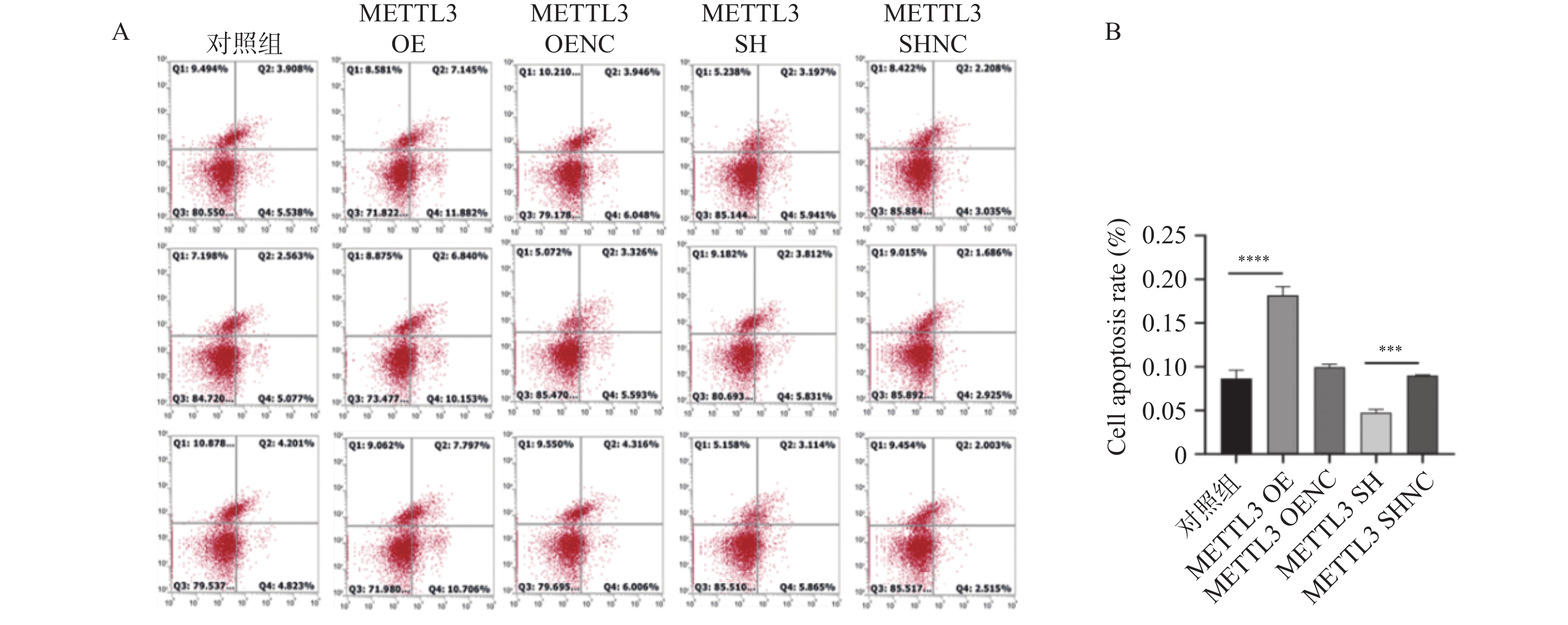
图 10 FITC/PI双染法检测颗粒细胞凋亡情况
A: FITC/PI双染法检测颗粒细胞凋亡情况 ;B:两组颗粒细胞凋亡结果。***P < 0.001,****P < 0.0001。
Figure 10. Detection of apoptosis of granulosa cells by FITC/PI double staining method
-
[1] Chon S J,Umair Z,Yoon M S. Premature ovarian insufficiency: Past,present,and future[J]. Frontiers in Cell and Developmental Biology,2021,9:672890. doi: 10.3389/fcell.2021.672890 [2] Tsiligiannis S,Panay N,Stevenson J C. Premature ovarian insufficiency and long-term health consequences[J]. Current Vascular Pharmacology,2019,17(6):604-609. doi: 10.2174/1570161117666190122101611 [3] E. F,Zhang H,Yin W,et al. CPEB3 deficiency in mice affect ovarian follicle development and causes premature ovarian insufficiency[J]. Cell Death & Disease,2021,13(1): 21. [4] Zhang J,Yan L,Wang Y,et al. In vivo and in vitro activation of dormant primordial follicles by EGF treatment in mouse and human[J]. Clinical and Translational Medicine,2020,10(5):e182. doi: 10.1002/ctm2.182 [5] Shao T,Ke H,Liu R,et al. Autophagy regulates differentiation of ovarian granulosa cells through degradation of WT1[J]. Autophagy,2022,18(8):1864-1878. doi: 10.1080/15548627.2021.2005415 [6] 潘正美,刘洋. 卵巢早衰小鼠模型综述[J]. Summary of Mouse Models of Premature Ovarian Failure,2021(12):97-102. [7] 李洁,杨菁,陈媛,等. D(+)半乳糖对小鼠卵巢功能影响的实验性研究[J]. 中国优生与遗传杂志,2006(12): 98-100+134. [8] Zhao Y T,Yin H,Hu C,et al. Tilapia skin peptides restore cyclophosphamide-induced premature ovarian failure via inhibiting oxidative stress and apoptosis in mice[J]. Food & Function,2022,13(3):1668-1679. [9] Rehnitz J,Youness B,Nguyen X P,et al. FMR1 expression in human granulosa cells and variable ovarian response: control by epigenetic mechanisms[J]. Molecular Human Reproduction,2021,27(2):gaab001. doi: 10.1093/molehr/gaab001 [10] 许梦婷,李芳,王莉莉. 卵巢功能早衰的病因与治疗研究进展[J]. 中国预防医学杂志,2022,23(5):394-400. [11] Zaccara S,Ries R J,Jaffrey S R. Reading,writing and erasing mRNA methylation[J]. Nature Reviews Molecular Cell Biology,2019,20(10):608-624. doi: 10.1038/s41580-019-0168-5 [12] Jiang X,Liu B,Nie Z,et al. The role of m6A modification in the biological functions and diseases[J]. Signal Transduction and Targeted Therapy,2021,6(1):74. doi: 10.1038/s41392-020-00450-x [13] Li G,Sun Z,Deng W,et al. METTL3 plays a crucial function in multiple biological processes[J]. Acta Histochemica,2022,124(6):151916. doi: 10.1016/j.acthis.2022.151916 [14] 杨智惠,胡扬,宗政,等. 卵巢功能不全病因病机及动物模型构建研究进展[J]. 中国比较医学杂志,2024,34(3):149-160. [15] Bhardwaj J K,Paliwal A,Saraf P,et al. Role of autophagy in follicular development and maintenance of primordial follicular pool in the ovary[J]. Journal of Cellular Physiology,2022,237(2):1157-1170. doi: 10.1002/jcp.30613 [16] Cakiroglu Y,Saltik A,Yuceturk A,et al. Effects of intraovarian injection of autologous platelet rich plasma on ovarian reserve and IVF outcome parameters in women with primary ovarian insufficiency[J]. Aging,2020,12(11):10211-10222. doi: 10.18632/aging.103403 [17] 刘艳艳,马园园,黄向华,等. 不同来源间充质干细胞治疗卵巢早衰的机制[J]. 中国组织工程研究,2025,29(13):2764-2773. [18] Jiao X,Zhang H,Ke H,et al. Premature Ovarian Insufficiency: Phenotypic Characterization Within Different Etiologies[J]. The Journal of Clinical Endocrinology & Metabolism,2017,102(7):2281-2290. [19] 江静,张明,陆嘉逢,等. RNA表观修饰在卵子发生调控中的研究进展[J]. 中国细胞生物学学报,2024,46(4):657-667. [20] Mu H,Zhang T,Yang Y,et al. METTL3-mediated mRNA N6-methyladenosine is required for oocyte and follicle development in mice[J]. Cell Death & Disease,2021,12(11):989. [21] Xia H,Zhong C,Wu X,et al. Mettl3 mutation disrupts gamete maturation and reduces fertility in zebrafish[J]. Genetics,2018,208(2):729-743. doi: 10.1534/genetics.117.300574 [22] Huang B,Ding C,Zou Q,et al. Cyclophosphamide regulates N6-methyladenosine and m6A RNA enzyme levels in human granulosa cells and in ovaries of a premature ovarian aging mouse model[J]. Frontiers in Endocrinology,2019,10:415. doi: 10.3389/fendo.2019.00415 [23] Li Z, Ruan Z, Feng Y, et al. METTL3-mediated m6A methylation regulates granulosa cells autophagy during follicular atresia in pig ovaries[J]. Theriogenology, 2023, 201: 83-94.Li Z,Ruan Z,Feng Y,et al. METTL3-mediated m6A methylation regulates granulosa cells autophagy during follicular atresia in pig ovaries[J]. Theriogenology,2023,201:83-94. -

 下载:
下载:

 点击查看大图
点击查看大图
计量
- 文章访问数: 395
- HTML全文浏览量: 299
- PDF下载量: 25
- 被引次数: 0
