

Correlation of MAPK1 and NRAS Gene Polymorphisms with Cervical Intraepithelial Neoplasia in Yunnan Han Population
-
摘要:
目的 探讨在云南汉族人群中NRAS基因与MAPK1基因rs14804和rs9340多态性位点与宫颈上皮内瘤变(cervical intraepithelial neoplasia,CIN)易感性的相关性。 方法 随机选取2017年5月至2019年10月昆明医科大学第三附属医院416例CIN患者和983例健康对照个体,通过TaqMan探针法对NRAS基因与MAPK1基因的SNPs位点(rs14804和rs9340)进行基因分型,分析2个SNPs位点与云南汉族人群CIN发生风险的相关性。 结果 MAPK1基因的SNP位点rs9340等位基因(P = 0.008)和基因型(P = 0.002)在CIN组与对照组的分布频率差异具有统计学意义,等位基因A可能与更高的CIN发生风险相关(OR = 1.28,95%CI 1.07 ~ 1.54),尤其是低年龄组(≤ 50岁)人群的CIN风险相关(OR = 1.35,95%CI 1.09 ~ 1.67)。 结论 MAPK1基因的SNP位点rs9340可能与云南汉族人群CIN发生风险具有相关性。 Abstract:Objective NRAS and MAPK1 genes are important regulators of the Ras/Raf/MEK/ERK signaling pathway, and their functions and expression levels have been associated with a variety of human tumors. This study aims to investigate the correlation between the rs14804 and rs9340 polymorphic loci of NRAS and MAPK1 genes and the susceptibility to cervical intraepithelial neoplasia (CIN) in Yunnan Han population. Methods 1399 persons of the population were enrolled in the current study, with 416 as the CIN patient group and 983 as the healthy control group. The SNPs loci (rs14804 and rs9340) in the NRAS gene and the MAPK1 gene were genotyped using the TaqMan assays, and the correlation between the two SNPs loci and the risk of CIN was analyzed. Results The difference in the frequency of distribution of SNP locus rs9340 allele (P = 0.008) and genotype (P = 0.002) of MAPK1 gene in the CIN group versus the control group was statistically significant, and allele A may be associated with a higher risk of developing CIN (OR = 1.28, 95%CI 1.07 ~ 1.54), especially for people in the lower age group (≤ 50 years) of CIN risk (OR = 1.35, 95%CI 1.09 ~ 1.67). Conclusion rs9340 in the MAPK1 gene might be correlated with the risk of CIN in the Chinese Han population in Yunnan. -
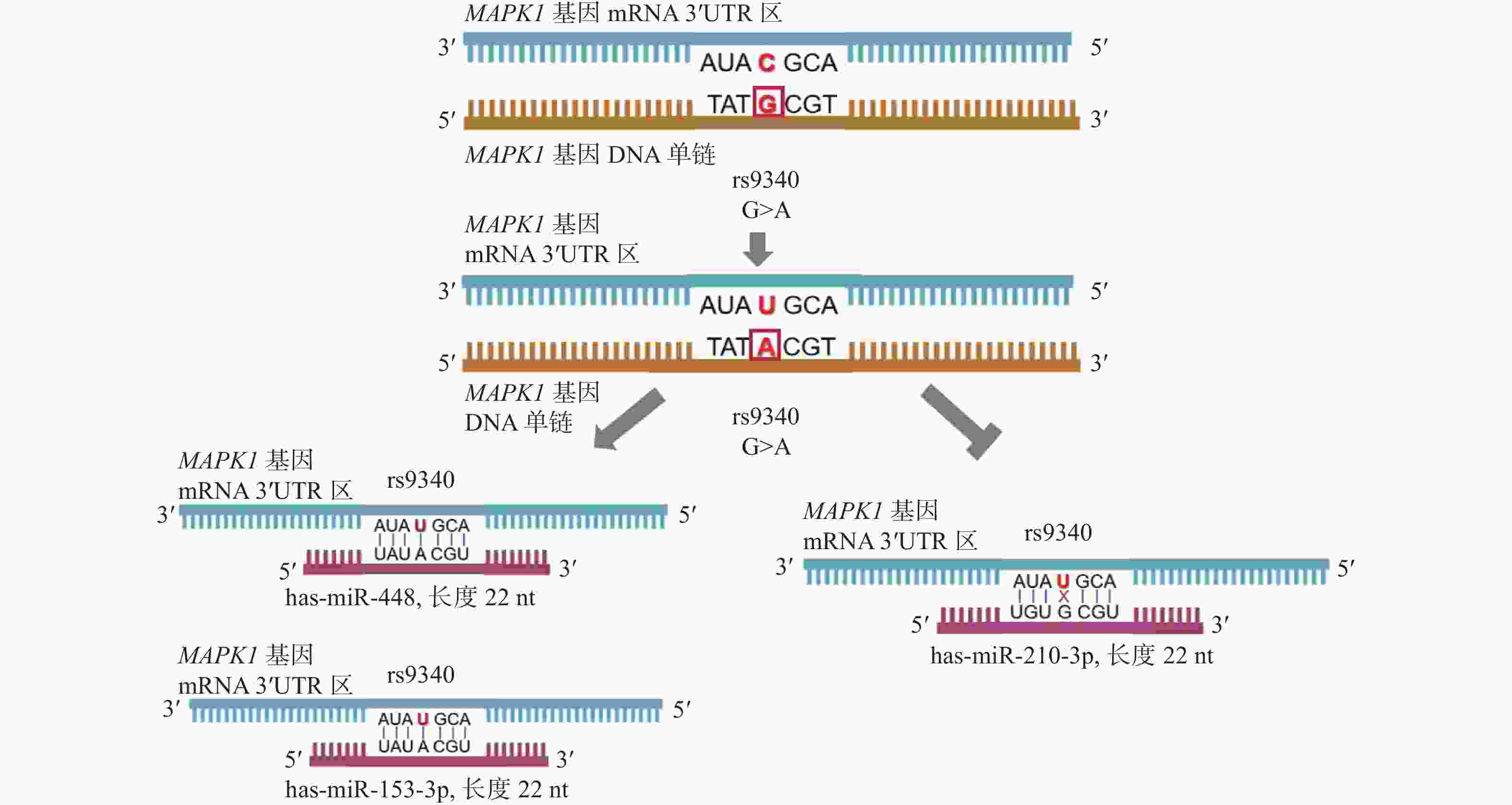
图 1 rs9340位点导致所在区域互补结合的miRNA发生改变
Figure 1. The rs9340 causes the different miRNA binding pattern
表 1 选取病例的临床特征[($ \bar{x} \pm s $),岁]
Table 1. The clinical characteristics of the subjects enrolled in this study [($ \bar{x} \pm s $),years old]
分组 临床分期/年龄分层 n 年龄分布 t P 对照组 高龄组 314 56.21 ± 3.85 低龄组 718 41.91 ± 6.86 总计 961 45.77 ± 8.87 CIN组 CIN2 51 45.59 ± 10.13 −1.535 0.903 CIN3 365 44.92 ± 9.48 −2.923 0.137 高龄组 102 58.09 ± 6.09 −3.523 0.00048* 低龄组 265 40.75 ± 5.92 −2.771 0.006* 总计 416 45.00 ± 9.55 −1.446 0.148 *P < 0.05。  下载: 导出CSV
下载: 导出CSV
表 2 2个SNPs位点在CIN和对照组间等位基因和基因型分布比较[n(%)]
Table 2. The comparison of allelic and genotypic distribution of the two SNPs between CIN and control groups[n(%)]
SNPs 等位基因/基因型 对照组 CIN组 χ2 P OR(95%CI) rs14804 A 49(2.50) 22(2.64) 0.055 0.815 1.06(0.64 ~ 1.77) G 1917(97.50) 810(97.36) A/A 0(0.00) 1(0.24) 2.382 0.304 A/G 49(4.98) 20(4.81) G/G 934(95.02) 395(94.95) HWE,P 0.423 0.177 rs9340 A 456(23.19) 232(27.88) 6.936 0.008* 1.28(1.07 ~ 1.54) G 1510(76.81) 600(72.12) A/A 55(5.60) 22(5.29) 12.568 0.002* A/G 346(35.20) 188(45.19) G/G 582(59.20) 206(49.52) HWE,P 0.705 0.012 *P < 0.025(统计学结果经Bonferroni校正,n = 2)。
下载: 导出CSV
表 3 高龄组和低龄组中rs9340位点与CIN的相关性分析[n(%)]
Table 3. The association of rs9340 with CIN in different age groups [n(%)]
年龄分层 等位基因/基因型 对照组 CIN组 χ2 P OR(95%CI) 高年龄组 A 131(24.72) 55(26.96) 0.108 0.743 1.07(0.73 ~ 1.55) G 399(75.28) 149(73.04) A/A 17(6.42) 4(3.92) 0.814 0.367 0.42(0.13 ~ 1.38) A/G 97(36.60) 47(46.08) 2.046 0.212 0.58(0.18 ~ 1.88) G/G 151(56.98) 51(50.00) 3.102 0.153 低年龄组 A 325(22.63) 177(28.18) 7.4 0.007* 1.35(1.09 ~ 1.67) G 1111(77.37) 451(71.82) A/A 38(5.29) 18(5.73) 10.37 0.001* 1.59(1.19 ~ 2.08) A/G 249(34.69) 141(44.90) 0.898 0.343 1.33(0.74 ~ 2.38) G/G 431(60.02) 155(49.37) 10.453 0.005 *P < 0.05。
下载: 导出CSV
表 4 rs9340位点与CIN分期进展的相关性[n(%)]
Table 4. Correlation of rs9340 locus polymorphism with different CIN stages [n(%)]
等位基因此/基因型 CIN2组 CIN3组 χ2 P OR(95%CI) A 22(21.57) 210(28.77) 2.323 0.127 0.68(0.41 ~ 1.12) G 80(78.43) 520(71.23) A/A 1(1.96) 21(5.75) 1.515 0.218 0.28(0.04 ~ 2.14) A/G 20(39.22) 168(46.03) 1.381 0.24 0.70(0.38~ 1.27) G/G 30(58.82) 176(48.22) 0b b该项为参照项,因此设置为0。
下载: 导出CSV
表 5 rs9340位点对MAPK1基因3'UTR区域miRNA互补结合的影响
Table 5. The effect of rs9340 on miRNA binding to the 3'UTR of MAPK1
下载: 导出CSV
-
[1] Bruni L A G,Serrano B,Mena M,et al. Human papillomavirus and related diseases in the world [R]. Barcelona,Spain,2023: ICO/IARC Information Centre on HPV and Cancer (HPV Information Centre),Summary Report 22 October 2021. [2] 冯明月,闫萍. 宫颈上皮内瘤变诊治现状[J]. 河北医科大学学报,2020,41(4):480-483. [3] Schiffman M, Castle P E, Jeronimo J, et al. Human papillomavirus and cervical cancer[J]. Lancet (London, England),2007,370(9590):890-907. [4] Chen D,Cui T,Ek W E,et al. Analysis of the genetic architecture of susceptibility to cervical cancer indicates that common SNPs explain a large proportion of the heritability[J]. Carcinogenesis,2015,36(9):992-998. doi: 10.1093/carcin/bgv083 [5] Vink J M,Van Kemenade F J,Meijer C J,et al. Cervix smear abnormalities: Linking pathology data in female twins,their mothers and sisters [J]. European Journal of Human Genetics : EJHG,2011,19(1): 108-111. [6] Zhang X,Zhang L,Tian C,et al. Genetic variants and risk of cervical cancer: Epidemiological evidence,meta-analysis and research review[J]. BJOG,2014,121(6):664-674. doi: 10.1111/1471-0528.12638 [7] Hatzivassiliou G,Song K,Yen I,et al. RAF inhibitors prime wild-type RAF to activate the MAPK pathway and enhance growth[J]. Nature,2010,464(7287):431-435. doi: 10.1038/nature08833 [8] Yuan J,Dong X,Yap J,et al. The MAPK and AMPK signalings: Interplay and implication in targeted cancer therapy[J]. Journal of Hematology & Oncology,2020,13(1):113. [9] Xie G,Zhu A,Gu X. Mitogen-activated protein kinase inhibition-induced modulation of epidermal growth factor receptor signaling in human head and neck squamous cell carcinoma[J]. Head & Neck,2021,43(6):1721-1729. [10] Zhou G,Yang J,Song P. Correlation of ERK/MAPK signaling pathway with proliferation and apoptosis of colon cancer cells[J]. Oncology Letters,2019,17(2):2266-2270. [11] Bartholomeusz C,Gonzalez-Angulo A M,Liu P,et al. High ERK protein expression levels correlate with shorter survival in triple-negative breast cancer patients[J]. The Oncologist,2012,17(6):766-774. doi: 10.1634/theoncologist.2011-0377 [12] Rauen K A. Defining RASopathy[J]. Dis Model Mech,2022,15(2):dmm049344. doi: 10.1242/dmm.049344 [13] Yan Y,Gao Z,Han H,et al. NRAS expression is associated with prognosis and tumor immune microenvironment in lung adenocarcinoma[J]. J Cancer Res Clin Oncol,2022,148(3):565-575. doi: 10.1007/s00432-021-03842-w [14] Wang Y,Guo Z,Tian Y,et al. MAPK1 promotes the metastasis and invasion of gastric cancer as a bidirectional transcription factor[J]. BMC Cancer,2023,23(1):959. doi: 10.1186/s12885-023-11480-3 [15] Harada G,Yang S R,Cocco E,et al. Rare molecular subtypes of lung cancer[J]. Nature Reviews Clinical Oncology,2023,20(4):229-249. doi: 10.1038/s41571-023-00733-6 [16] Cicenas J,Tamosaitis L,Kvederaviciute K,et al. KRAS,NRAS and BRAF mutations in colorectal cancer and melanoma[J]. Medical Oncology (Northwood,London,England),2017,34(2):26. doi: 10.1007/s12032-016-0879-9 [17] Ekedahl H, Cirenajwis H, Harbst K, et al. The clinical significance of BRAF and NRAS mutations in a clinic-based metastatic melanoma cohort[J]. Br J Dermatol,2013,169(5):1049-1055. [18] Emrick M A,Hoofnagle A N,Miller A S,et al. Constitutive activation of extracellular signal-regulated kinase 2 by synergistic point mutations[J]. The Journal of Biological Chemistry,2001,276(49):46469-46479. doi: 10.1074/jbc.M107708200 [19] Ojesina A I,Lichtenstein L,Freeman S S,et al. Landscape of genomic alterations in cervical carcinomas[J]. Nature,2014,506(7488):371-375. doi: 10.1038/nature12881 [20] Insodaite R,Smalinskiene A,Liutkevicius V,et al. Associations of polymorphisms localized in the 3'UTR regions of the KRAS,NRAS,MAPK1 genes with laryngeal squamous cell carcinoma[J]. Genes,2021,12(11):1679. doi: 10.3390/genes12111679 [21] 国家癌症中心,国家肿瘤质控中心宫颈癌质控专家委员会. 中国宫颈癌规范诊疗质量控制指标(2022版)[J]. 中华肿瘤杂志,2022,44(7):615-622. [22] 中华人民共和国国家卫生和计划生育委员会. 宫颈癌及癌前病变规范化诊疗指南(试行)[J]. 中国医学前沿杂志(电子版),2013,5(8):40-49. [23] Bhatla N,Aoki D,Sharma D N,et al. Cancer of the cervix uteri[J]. International Journal of Gynecology & amp; Obstetrics,2018,143(S2):22-36. [24] 中华医学会妇产科学分会绝经学组. 中国绝经管理与绝经激素治疗指南2023版[J]. 中华妇产科杂志,2023,58(1):4-21. [25] Ma H,Tian T,Liu X,et al. Upregulated circ_0005576 facilitates cervical cancer progression via the miR-153/KIF20A axis[J]. Biomed Pharmacother,2019(10):109311. doi: 10.1016/j.biopha.2019.109311 [26] Shao M X,Qu A Z,Wang Y Q,et al. Expression level of miRNA-210-3p in cervical cancer and its prognostic potential[J]. Eur Rev Med Pharmacol Sci,2020,24(12):6583-6588. [27] Bos J L. The ras gene family and human carcinogenesis[J]. Mutation Research,1988,195(3):255-271. doi: 10.1016/0165-1110(88)90004-8 [28] Guin S,Theodorescu D. The RAS-RAL axis in cancer: Evidence for mutation-specific selectivity in non-small cell lung cancer[J]. Acta Pharmacologica Sinica,2015,36(3):291-297. doi: 10.1038/aps.2014.129 [29] Lanfredini S,Thapa A,O'neill E. RAS in pancreatic cancer[J]. Biochemical Society Transactions,2019,47(4):961-972. doi: 10.1042/BST20170521 [30] Afrăsânie V A,Marinca M V,Alexa-Stratulat T,et al. KRAS,NRAS,BRAF,HER2 and microsatellite instability in metastatic colorectal cancer - practical implications for the clinician[J]. Radiology and Oncology,2019,53(3):265-274. doi: 10.2478/raon-2019-0033 [31] Li S,Balmain A,Counter C M. A model for RAS mutation patterns in cancers: Finding the sweet spot[J]. Nature Reviews Cancer,2018,18(12):767-777. doi: 10.1038/s41568-018-0076-6 [32] Jakob J A,Bassett R L J r,Ng C S,et al. NRAS mutation status is an independent prognostic factor in metastatic melanoma[J]. Cancer,2012,118(16):4014-4023. doi: 10.1002/cncr.26724 [33] Schirripa M,Cremolini C,Loupakis F,et al. Role of NRAS mutations as prognostic and predictive markers in metastatic colorectal cancer[J]. International Journal of Cancer,2015,136(1):83-90. doi: 10.1002/ijc.28955 [34] Murphy B M, Terrell E M, Chirasani V R, et al. Enhanced BRAF engagement by NRAS mutants capable of promoting melanoma initiation[J]. Nat Commun,2022,13(1):3153. [35] Fu W,Zhuo Z,Hua R X,et al. Association of KRAS and NRAS gene polymorphisms with Wilms tumor risk: A four-center case-control study[J]. Aging,2019,11(5):1551-1563. doi: 10.18632/aging.101855 [36] Alessandro L,Low K E,Abushelaibi A,et al. Identification of NRAS diagnostic biomarkers and drug targets for endometrial cancer-an integrated in silico approach[J]. International Journal of Molecular Sciences,2022,23(22):14285. doi: 10.3390/ijms232214285 [37] Jin M,Li Z,Sun Y,et al. Association analysis between the interaction of RAS family genes mutations and papillary thyroid carcinoma in the Han Chinese population[J]. International Journal of Medical Sciences,2021,18(2):441-447. doi: 10.7150/ijms.50026 [38] Li S,Ma Y M,Zheng P S,et al. GDF15 promotes the proliferation of cervical cancer cells by phosphorylating AKT1 and Erk1/2 through the receptor ErbB2 [J]. Journal of Experimental & Clinical Cancer Research : CR,2018,37(1): 80. [39] Yan Z,Ohuchida K,Fei S,et al. Inhibition of ERK1/2 in cancer-associated pancreatic stellate cells suppresses cancer-stromal interaction and metastasis [J]. Journal of Experimental & Clinical Cancer Research : CR,2019,38(1): 221. [40] Deng R,Zhang H L,Huang J H,et al. MAPK1/3 kinase-dependent ULK1 degradation attenuates mitophagy and promotes breast cancer bone metastasis[J]. Autophagy,2021,17(10):3011-3029. doi: 10.1080/15548627.2020.1850609 [41] Wills C, Watts K, Maughan T S, et al. Germline variation in RASAL2 may predict survival in patients with RAS-activated colorectal cancer[J]. Genes Chromosomes Cancer,2023,62(6):332-341. [42] Campbell J D,Alexandrov A,Kim J,et al. Distinct patterns of somatic genome alterations in lung adenocarcinomas and squamous cell carcinomas[J]. Nature Genetics,2016,48(6):607-616. doi: 10.1038/ng.3564 [43] Santos M,Lima L,Carvalho S,et al. The impact of BDNF,NTRK2,NGFR,CREB1,GSK3B,AKT,MAPK1,MTOR,PTEN,ARC,and SYN1 genetic polymorphisms in antidepressant treatment response phenotypes[J]. International Journal of Molecular Sciences,2023,24(7):6758. doi: 10.3390/ijms24076758 [44] Zhu Y, Yang T, Duan J, et al. MALAT1/miR-15b-5p/MAPK1 mediates endothelial progenitor cells autophagy and affects coronary atherosclerotic heart disease via mTOR signaling pathway[J]. Aging,2019,11(4):1089-1109. [45] Guney G,Taşkın M I,Sener N,et al. The role of ERK-1 and ERK-2 gene polymorphisms in PCOS pathogenesis [J]. Reproductive Biology and Endocrinology : RB&E,2022,20(1): 95. [46] Wei H,Ke H L,Lin J,et al. MicroRNA target site polymorphisms in the VHL-HIF1α pathway predict renal cell carcinoma risk[J]. Molecular Carcinogenesis,2014,53(1):1-7. doi: 10.1002/mc.21917 [47] Guo N,Zhang N,Yan L,et al. Correlation between genetic polymorphisms within the MAPK1/HIF-1/HO-1 signaling pathway and risk or prognosis of perimenopausal coronary artery disease[J]. Clinical Cardiology,2017,40(8):597-604. doi: 10.1002/clc.22708 -

 点击查看大图
点击查看大图

计量
- 文章访问数: 2934
- HTML全文浏览量: 1069
- PDF下载量: 24
- 被引次数: 0
