

miR-125b-5p Regulates HK2 to Inhibit Proliferation and Glycolysis of Gallbladder Cancer Cells
-
摘要:
目的 研究miR-125b-5p通过HK2调控人胆囊癌细胞增殖和糖酵解的作用机制。 方法 分别在人胆囊癌细胞GBC-SD中转染NC mimic、miR-125b-5p mimic、miR-125b-5p mimic+pcDNA-NC、miR-125b-5p mimic+pcDNA-HK2。实时荧光定量聚合酶链式反应(Real Time Quantitative Polymerase Chain Reaction,RT-qPCR)检测miR-125b-5p和HK2 mRNA的表达,Western blot检测HK2和LDHA、PDK1的蛋白表达。细胞计数试剂盒-8(cell countingKit-8,CCK-8)检测细胞增殖活力,糖酵解相关试剂盒检测乳酸生成、葡萄糖消耗以及ATP生成。双萤光素酶报告基因实验对miR-125b-5p和HK2的靶向关系进行验证。 结果 miR-125b-5p在胆囊癌细胞系GBC-SD和NOZ(P < 0.001)以及组织(P < 0.01)中表达降低,且在GBC-SD细胞中低于NOZ细胞中。过表达miR-125b-5p可显著抑制GBC-SD细胞的增殖(P < 0.05)、葡萄糖消耗(P < 0.05)、乳酸(P < 0.001)和ATP的生成(P < 0.01)以及LDHA(P < 0.01)、PDK1(P < 0.001)、HK2(P < 0.0001)表达。starBase数据库和双萤光素酶报告基因实验证实miR-125b-5p靶向负调控HK2。过表达miR-125b-5p和HK2组中细胞的增殖(P < 0.01)、葡萄糖摄取(P < 0.05)、乳酸(P < 0.01)和ATP(P < 0.05)的生成以及LDHA(P < 0.05)、PDK1(P < 0.05)、HK2(P < 0.05)表达显著高于仅过表达miR-125b-5p组。 结论 过表达miR-125b-5p靶向HK2,抑制人胆囊癌细胞GBC-SD的增殖和有氧糖酵解。 -
关键词:
- 胆囊癌 /
- miR-125b-5p /
- 增殖 /
- 糖酵解 /
- HK2
Abstract:Objective To investigate the mechanism of miR-125b-5p regulating the proliferation and glycolysis of gallbladder cancer cells by HK2. Methods NC mimic, miR-125b-5p mimic, miR-125b-5p mimic+pcDNA-NC and miR-125b-5p mimic+pcDNA-HK2 were transfected into human gallbladder carcinoma GBC-SD cells, respectively. Real Time Quantitative Polymerase Chain Reaction (RT-qPCR) was used to detect the expression of miR-125b-5p and HK2 mRNA. The protein expressions of HK2, LDHA and PDK1 were detected by Western blot. Cell counting Kit-8 (CCK-8) was used to measure cell proliferation activity, and the glycolysis kit was used to measure lactic acid production, glucose consumption, and ATP production. Dual luciferase reporter assay was used to verify the targeting relationship between miR-125b-5p and HK2. Results The expression of miR-125b-5p was reduced in gallbladder carcinoma cell lines GBC-SD, NOZ (P < 0.0001) and tissues (P < 0.01), and was lower in GBC-SD cells than in NOZ cells. Overexpression of miR-125b-5p significantly inhibited the proliferation of GBC-SD cells (P < 0.05), glucose intake (P < 0.05), the production of lactic acid (P < 0.001) and ATP production (P < 0.01), and the expression of LDHA (P < 0.01), PDK1 (P < 0.001), and HK2 (P < 0.0001). starBase database and dual luciferase experiments confirmed that miR-125b-5p targeted HK2 and negatively regulated its expression. Cell proliferation rate (P < 0.01), glucose consumption (P < 0.05), lactic acid (P < 0.01) and ATP production (P < 0.05), and the expression of LDHA (P < 0.05), PDK1 ( P < 0.05)、HK2 (P < 0.05)were significantly higher in the miR-125b-5p and HK2 overexpression groups. Conclusion Overexpression of miR-125b-5p inhibited the proliferation and aerobic glycolysis in human gallbladder carcinoma cell GBC-SD through targeting HK2. -
Key words:
- Gallbladder cancer /
- miR-125b-5p /
- Proliferation /
- Glycolysis /
- HK2
-

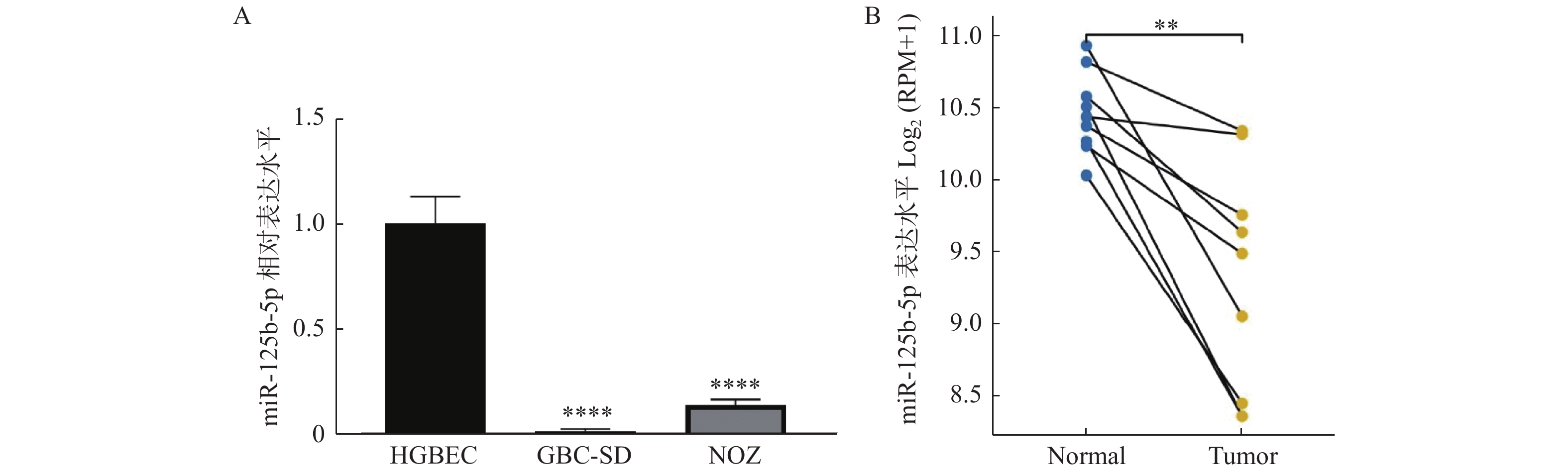
图 1 miR-125b-5p在胆囊癌细胞和组织中表达降低
A:RT-qPCR检测miR-125b-5p的表达,与HGBEC组相比,****P < 0.0001;B:TCGA数据库预测miR-125b-5p胆囊癌组织和癌旁组织中的表达,与Normal组相比,**P < 0.01。
Figure 1. The expression of miR-125b-5p was decreased ingallbladder carcinoma cells and tissues

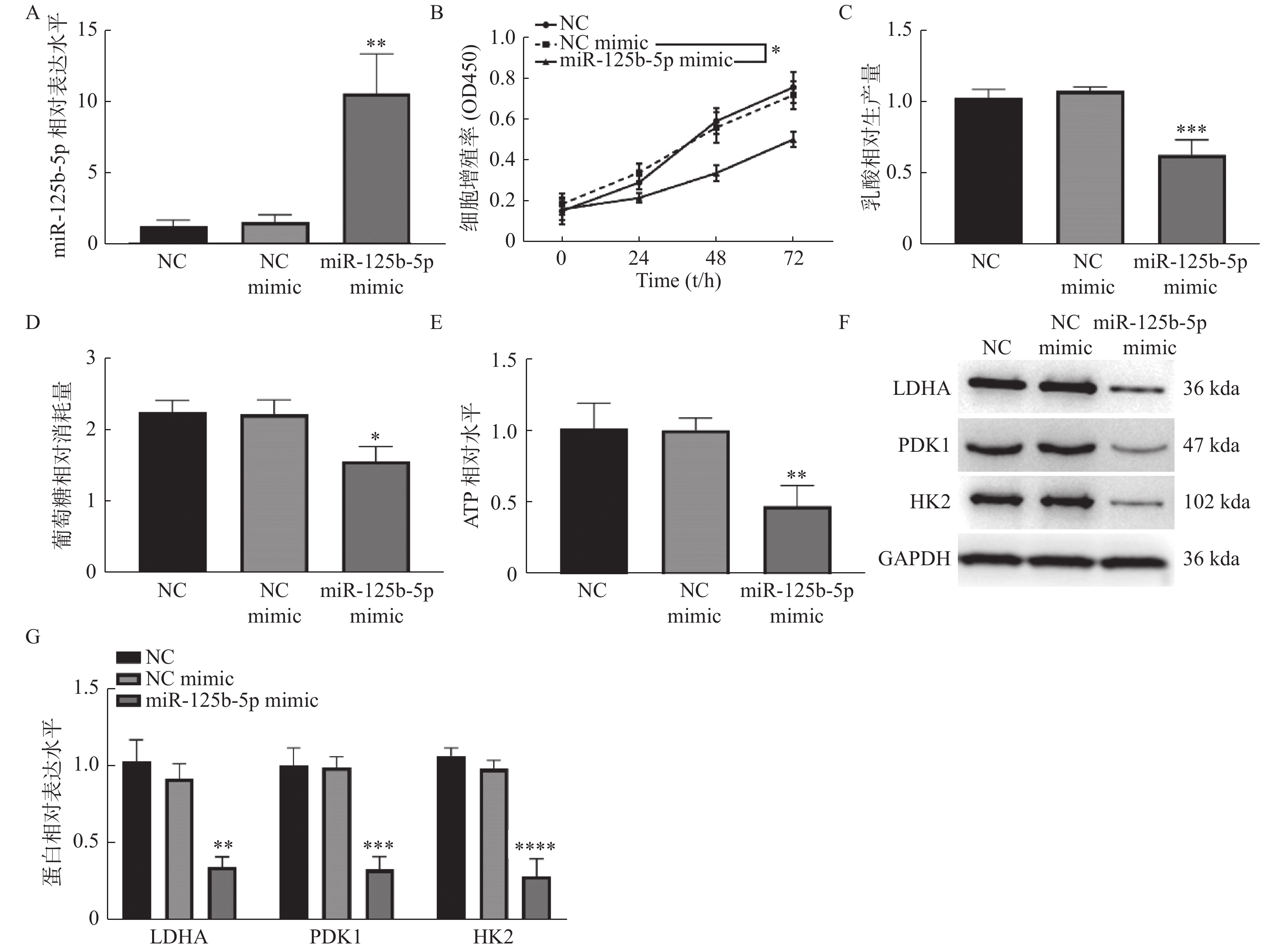
图 2 过表达miR-125b-5p抑制GBC-SD细胞增殖和有氧糖酵解
A:RT-qPCR检测miR-125b-5pmimic的转染效率;B:CCK-8检测细胞增殖活力;C:乳酸生成量检测;D:葡萄糖消耗量检测;E:ATP水平检测;F:Western blot检测LDHA、PDK1和的蛋白表达。HK2与NC mimic组相比,*P < 0.05,**P < 0.01,***P < 0.001,****P < 0.0001。
Figure 2. Over-expression of miR-125b-5p inhibited the proliferation and aerobic glycolysis in GBC-SD cell

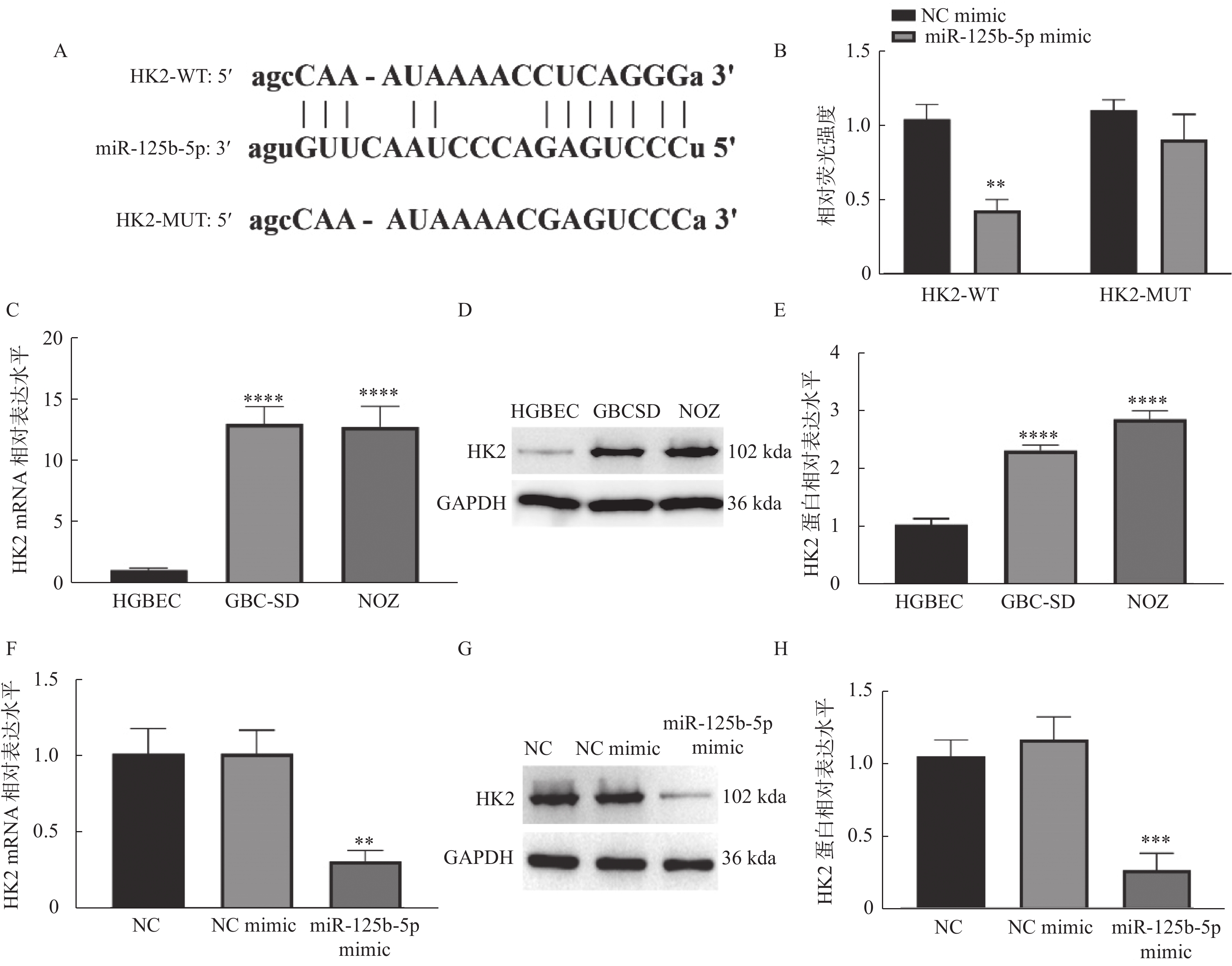
图 3 miR-125b-5p靶向负调控HK2
A:starBase数据数据库预测miR-125b-5p和HK2的靶向结合位点;B:双荧光素酶实验验证靶向关系,与NC mimic组相比,**P < 0.01;C:RT-qPCR检测HK2在胆囊癌和胆囊上皮细胞系中的mRNA水平,与HGBEC组相比,****P < 0.0001;D:Western blot检测HK2在胆囊癌和胆囊上皮细胞系中的蛋白水平,与HGBEC组相比,****P < 0.0001;E:RT-qPCR检测过表达miR-125-5p对HK2 mRNA水平的影响,与NC mimic组相比,**P < 0.01;F:Western blot检测过表达miR-125-5p对HK2蛋白水平的影响,与NC mimic组相比,***P < 0.001;G-H:Western blot检测过表达miR-125-5p对HK2蛋白水平的影响,与NC mimic组相比,***P < 0.001。
Figure 3. miR-125b-5p targeted and negatively regulated HK2

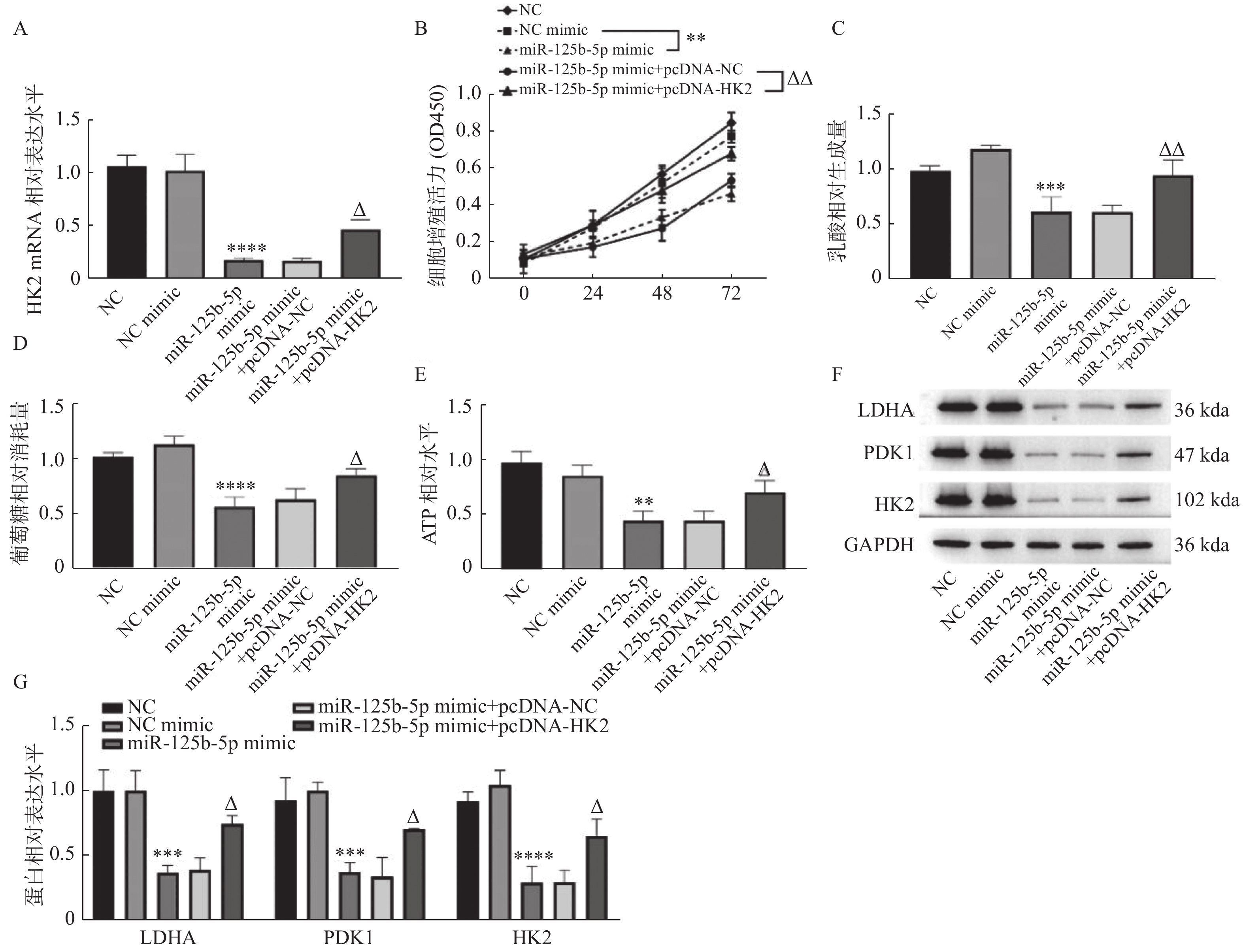
图 4 miR-125b-5p靶向HK2抑制GBC-SD细胞增殖和有氧糖酵解
A:RT-qPCR检测HK2 mRNA表达;B:CCK-8检测细胞增殖活力;C:乳酸含量检测;D:葡萄糖消耗量检测;E:ATP水平检测;F和G:LDHA、PDK1和HK2的蛋白水平测定。与NC mimic组相比,**P < 0.01,***P < 0.001,****P < 0.0001;与miR-125b-5p mimic+pcDNA-NC组相比,△P < 0.05,△△P < 0.01。
Figure 4. miR-125b-5p inhibited theproliferationandaerobic glycolysis in GBC-SD cellby targeting HK2
表 1 引物序列
Table 1. Primer sequences
基因名 引物序列 (F:上游引物,R:下游引物,5′-3′) miR-125b-3p F:TCCCTGAGACCCTAACTTGTGA R:TCACAAGTTAGGGTCTCAGGGA U6 F:CGCTTCGGCAGCACATATAC R:AATATGGAACGCTTCACGA  下载: 导出CSV
下载: 导出CSV
-
[1] Sharma A,Sharma K L,Gupta A,et al. Gallbladder cancer epidemiology,pathogenesis and molecular genetics:Recent update[J]. World J Gastroenterol,2017,23(22):3978-3998. doi: 10.3748/wjg.v23.i22.3978 [2] Gong Y Q,Ni J L,Fang Q,et al. MiR-1231 enhances docetaxel sensitivity to gallbladder carcinoma cells by downregulating FOXC2[J]. Eur Rev Med Pharmacol Sci,2020,24(23):12116-12123. [3] Zheng Y,Liu P,Wang N,et al. Betulinic acid suppresses breast cancer metastasis by targeting GRP78-mediated glycolysis and ER stress apoptotic pathway[J]. Oxid Med Cell Longev,2019,2019:8781690. [4] Li J,Hu Z Q,Yu S Y,et al. CircRPN2 inhibits aerobic glycolysis and metastasis in hepatocellular carcinoma[J]. Cancer Res,2022,82(6):1055-1069. doi: 10.1158/0008-5472.CAN-21-1259 [5] Arora S,Singh P,Tabassum G,et al. miR-16-5p regulates aerobic glycolysis and tumorigenesis of NSCLC cells via LDH-A/lactate/NF-κB signaling[J]. Life Sci,2022,304:120722. doi: 10.1016/j.lfs.2022.120722 [6] Zhu Y,Li F,Wan Y,et al. Cancer-secreted exosomal miR-620 inhibits ESCC aerobic glycolysis via FOXM1/HER2 pathway and promotes metastasis[J]. Front Oncol,2022,12:756109. doi: 10.3389/fonc.2022.756109 [7] Ren J,Li W,Pan G,et al. miR-142-3p modulates cell invasion and migration via PKM2-mediated aerobic glycolysis in colorectal cancer[J]. Anal Cell Pathol (Amst),2021,2021:9927720. [8] Zhai D,Li T,Ye R,et al. LncRNA LGALS8-AS1 promotes breast cancer metastasis through miR-125b-5p/SOX12 feedback regulatory network[J]. Front Oncol,2021,11:711684. doi: 10.3389/fonc.2021.711684 [9] Mei L L,Wang W J,Qiu Y T,et al. miR-125b-5p functions as a tumor suppressor gene partially by regulating HMGA2 in esophageal squamous cell carcinoma[J]. PLoS One,2017,12(10):e0185636. doi: 10.1371/journal.pone.0185636 [10] Hu B,Yang X B,Yang X,et al. LncRNA CYTOR affects the proliferation,cell cycle and apoptosis of hepatocellular carcinoma cells by regulating the miR-125b-5p/KIAA1522 axis[J]. Aging (Albany NY),2020,13(2):2626-2639. [11] Hua Q,Jin M,Mi B,et al. LINC01123,a c-Myc-activated long non-coding RNA,promotes proliferation and aerobic glycolysis of non-small cell lung cancer through miR-199a-5p/c-Myc axis[J]. J Hematol Oncol,2019,12(1):91. doi: 10.1186/s13045-019-0773-y [12] Randi G,Franceschi S,La Vecchia C. Gallbladder cancer worldwide:Geographical distribution and risk factors[J]. Int J Cancer,2006,118(7):1591-602. doi: 10.1002/ijc.21683 [13] Sharma Y,Saini A K,Kashyap S,et al. Host miRNA and immune cell interactions:Relevance in nano-therapeutics for human health[J]. Immunol Res,2022,70(1):1-18. doi: 10.1007/s12026-021-09247-8 [14] Jin F E,Xie B,Xian H Z,et al. Knockdown of miR-125b-5p inhibits the proliferation and invasion of gastric carcinoma cells by targeting RYBP[J]. Kaohsiung J Med Sci,2021,37(10):863-871. doi: 10.1002/kjm2.12425 [15] Cao J Y,Wang B,Tang T T,et al. Exosomal miR-125b-5p deriving from mesenchymal stem cells promotes tubular repair by suppression of p53 in ischemic acute kidney injury[J]. Theranostics,2021,11(11):5248-5266. doi: 10.7150/thno.54550 [16] Yang D,Zhan M,Chen T,et al. miR-125b-5p enhances chemotherapy sensitivity to cisplatin by down-regulating Bcl2 in gallbladder cancer[J]. Sci Rep,2017,7:43109. doi: 10.1038/srep43109 [17] Wang J,Huang Q,Hu X,et al. Disrupting circadian rhythm via the PER1-HK2 axis reverses trastuzumab resistance in gastric cancer[J]. Cancer Res,2022,82(8):1503-1517. [18] Guo D,Tong Y,Jiang X,et al. Aerobic glycolysis promotes tumor immune evasion by hexokinase2-mediated phosphorylation of IκBα[J]. Cell Metab,2022,34(9):1312-1324.e6. doi: 10.1016/j.cmet.2022.08.002 [19] Shi T,Ma Y,Cao L,et al. B7-H3 promotes aerobic glycolysis and chemoresistance in colorectal cancer cells by regulating HK2[J]. Cell Death Dis,2019,10(4):308. [20] Lu J, Zhen S, Tuo X, et al. Downregulation of DNMT3A attenuates the warburg effect, proliferation, and invasion via promoting the inhibition of miR-603 on HK2 in ovarian cancer[J]. Technol Cancer Res Treat, 2022, 21: 15330338221110668. [21] Xu Q L,Luo Z,Zhang B,et al. Methylation-associated silencing of miR-9-1 promotes nasopharyngeal carcinoma progression and glycolysis via HK2[J]. Cancer Sci,2021,112(10):4127-4138. doi: 10.1111/cas.15103 [22] Chen J,Yu Y,Li H,et al. Long non-coding RNA PVT1 promotes tumor progression by regulating the miR-143/HK2 axis in gallbladder cancer[J]. Mol Cancer,2019,18(1):33. doi: 10.1186/s12943-019-0947-9 -

 点击查看大图
点击查看大图
计量
- 文章访问数: 4637
- HTML全文浏览量: 2340
- PDF下载量: 37
- 被引次数: 0
