

Metagenomic Analysis of Gut Microbiome in Infants with or without Breast Milk Jaundice
-
摘要:
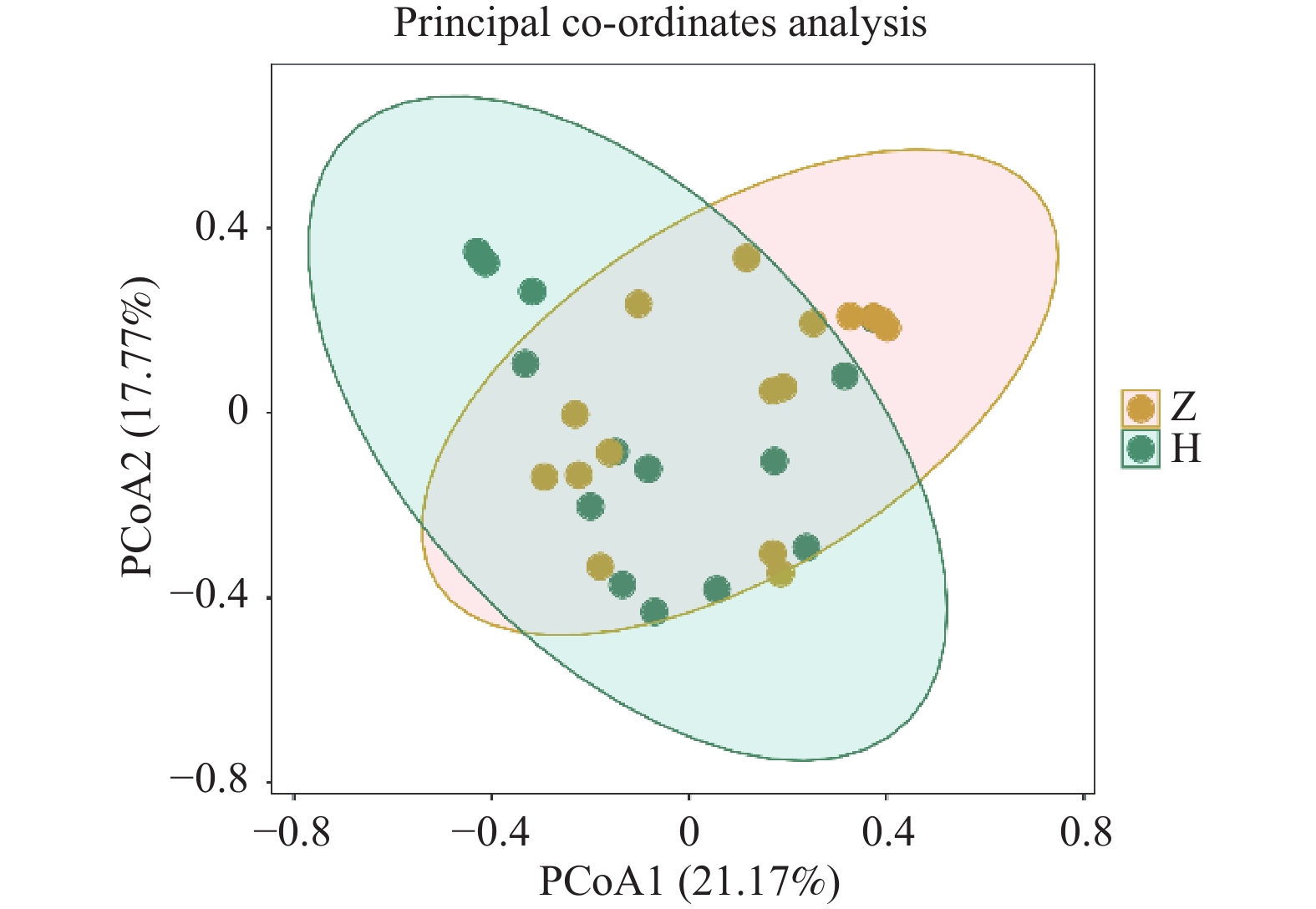
目的 探讨纯母乳喂养方式下,出现迟发型母乳性黄疸(late-onset breast milk jaundice,LBMJ)及未出现黄疸的婴儿之间肠道菌群的差异,以及可能影响 LBMJ 发生和发展的差异菌群及代谢途径。 方法 选取健康母婴16对,LBMJ 的母婴15对,月龄在15d至3月,满足足月且纯母乳喂养,分别命名为正常组(Z组)、黄疸组(H组),无菌取粪便样本随后冷冻保存。采两组肠道微生物的差异性通过宏基因组学的方法进行检测和分析。组间多样性分析应用 Kruskal - Wallis 检验,差异性分析采用Mann Whitney U检验等。 结果 所有样本肠道测序序列共注释到53个菌门, 1028 个菌属,5421 个菌种。H组的 Alpha多样性与Z组相比降低(P < 0.05),Beta多样性则没有显著的区别(P>0.05)。两组间肠道菌群之间的物种差异分析发现,Z组在门水平富集了衣原体,Deinococcota,Cyanobacteriota,Spirochaetota;在属水平,Z组中志贺杆菌,Eggerthella等明显增加,H组中葡萄球菌,Atopobium有明显增加;在种水平,志贺杆菌在Z组中显著增加,而未分类的葡萄球菌在H组中显著增加。LEfSe差异分析,Z组标志物为Coriobacteriia,Hungatella,Enterocloster,Flavonifractor,志贺杆菌属,Clostridioides,Flavonifractor plautii,Enterocloster bolteae,Enterocloster unclassified,宋内志贺菌等,H标志物为芽孢杆菌目,葡萄球菌科,拟杆菌噬菌体等。GO功能富集预测结果显示,Z组的细胞组分中的细胞质膜、细胞质及分子功能中蛋白质结合途径显著富集,H组在合成UDP−葡萄糖醛酸的生过程显著富集。KEGG功能预测发现,Z组在第三层级中对于二恶英降解、耶尔森氏鼠疫杆菌感染通路具有优势,而H组在次灵杆菌素生物合成通路方面占优势。第四层级,在与生物合成代谢相关均在Z组更具优势。结论 纯母乳喂养的 LBMJ 婴儿与非黄疸婴儿肠道菌群物种组成、标志物及基因功能注释存在显著差异,可能会对LBMJ发生和发展造成影响。 Abstract:Objective To explore the differences in gut microbiota composition of exclusively breastfed infants with and without late-onset breast milk jaundice (LBMJ), and to identify key bacterial taxa and metabolic pathways associated with LBMJ development and progression. Methods We selected 16 healthy full-term mother-infant pairs with exclusively breastfed infants and 15 healthy full-term mother-infant pairs diagnosed with late-onset breast milk jaundice, both groups with infants aged 0.5 to 3 months. The healthy exclusively breastfed infants were assigned to the control group (Group Z), and those with late-onset breast milk jaundice to the jaundice group (Group H). Fecal samples were collected under aseptic conditions and preserved at −80 °C. Differences in intestinal microbiota between the two groups were detected and analyzed using metagenomic methods. Inter-group diversity analysis was performed using the Kruskal-Wallis test, and differential analysis employed the Mann-Whitney U test, among others. Results All fecal microbial sequences were annotated into 53 phyla, 1028 genera, and5421 species. Alpha diversity in the H group was significantly reduced compared to the Z group (P<0.05), while Beta diversity showed no significant difference (P < 0.05). Species-level differential analysis between the two groups revealed that at the phylum level, group Z showed enrichment of Chlamydia, Deinococcota, Cyanobacteriota, and Spirochaetota. At the genus level, Shigella and Eggerthella were significantly increased in the Z group, while Staphylococcus and Atopobium were significantly increased in the H group. At the species level, Shigella was significantly elevated in the Z group, while unclassified Staphylococcus was significantly increased in the H group. LEfSe differential analysis revealed biomarkers for the Z group including Coriobacteriia, Hungatella, Enterocloster, Flavonifractor, Shigella, Clostridioides, Flavonifractor plautii, Enterocloster bolteae, Enterocloster unclassified, Shigella sonnei, while H group biomarkers included Bacillales, Staphylococcaceae, and Bacteriophage. Gene Ontology functional enrichment predictions showed that cellular components including cytoplasmic membrane and cytoplasm, as well as molecular functions including protein binding pathways, were significantly enriched in the Z group. The H group showed significant enrichment in UDP-glucuronic acid synthesis pathways. KEGG function predictions revealed that the Z group demonstrated advantages in dioxin degradation and Yersinia pestis infection pathways at the tertiary level, while the H group showed advantages in bacitracin biosynthesis pathways. At the quaternary level, biosynthetic metabolism-related pathways were more advantageous in the Z group.Conclusions Exclusively breastfed infants with LBMJ show significant differences from non-jaundiced infants in intestinal microbiota species composition, biomarkers, and gene function annotation, which may influence the occurrence and development of LBMJ. -
Key words:
- Late-onset breast milk jaundice /
- Exclusive breastfeeding /
- Gut microbiota /
- Metagenomics
-
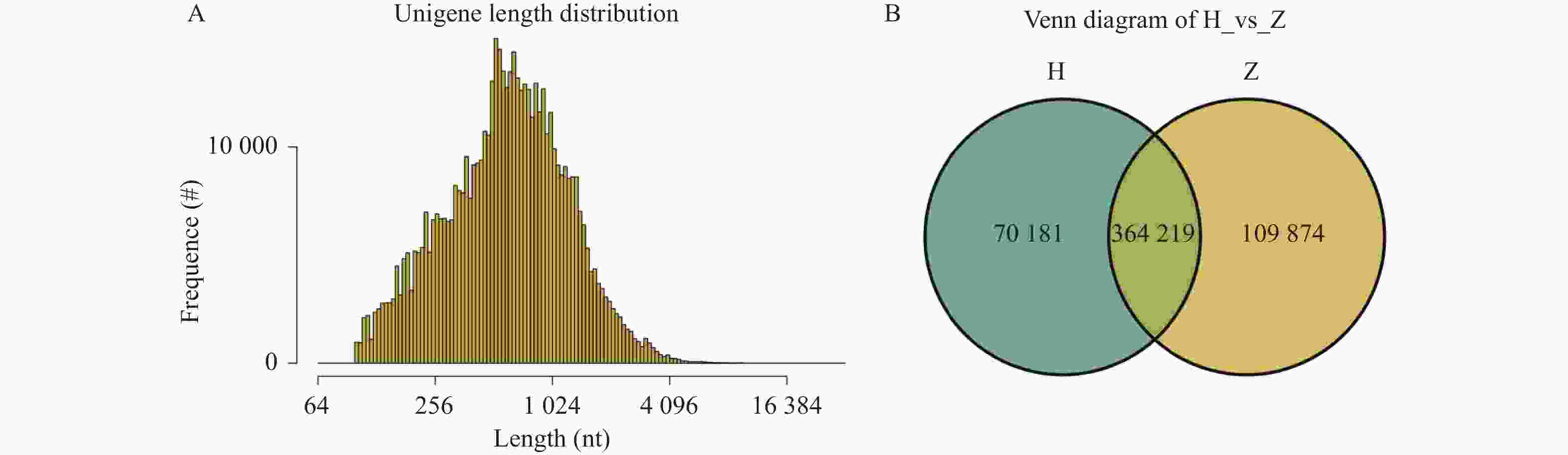
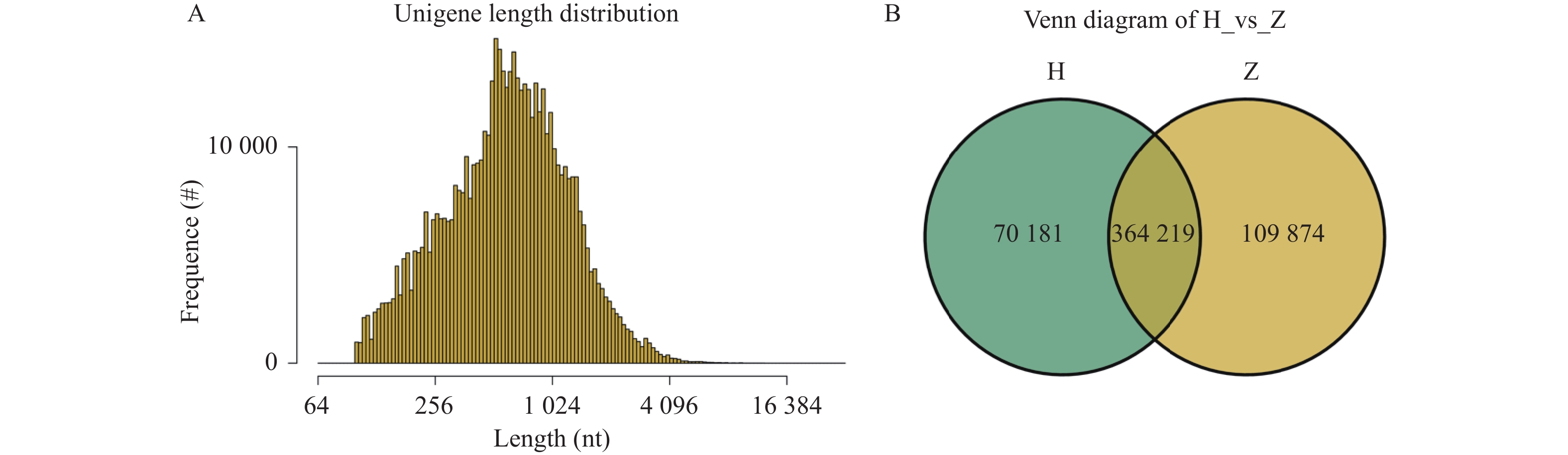
图 2 基因长度分布图和组间基因数目Venn图
A:基因长度分布图;B:组间基因数目Venn图。
Figure 2. Gene length distribution and venn plot of gene numbers among groups
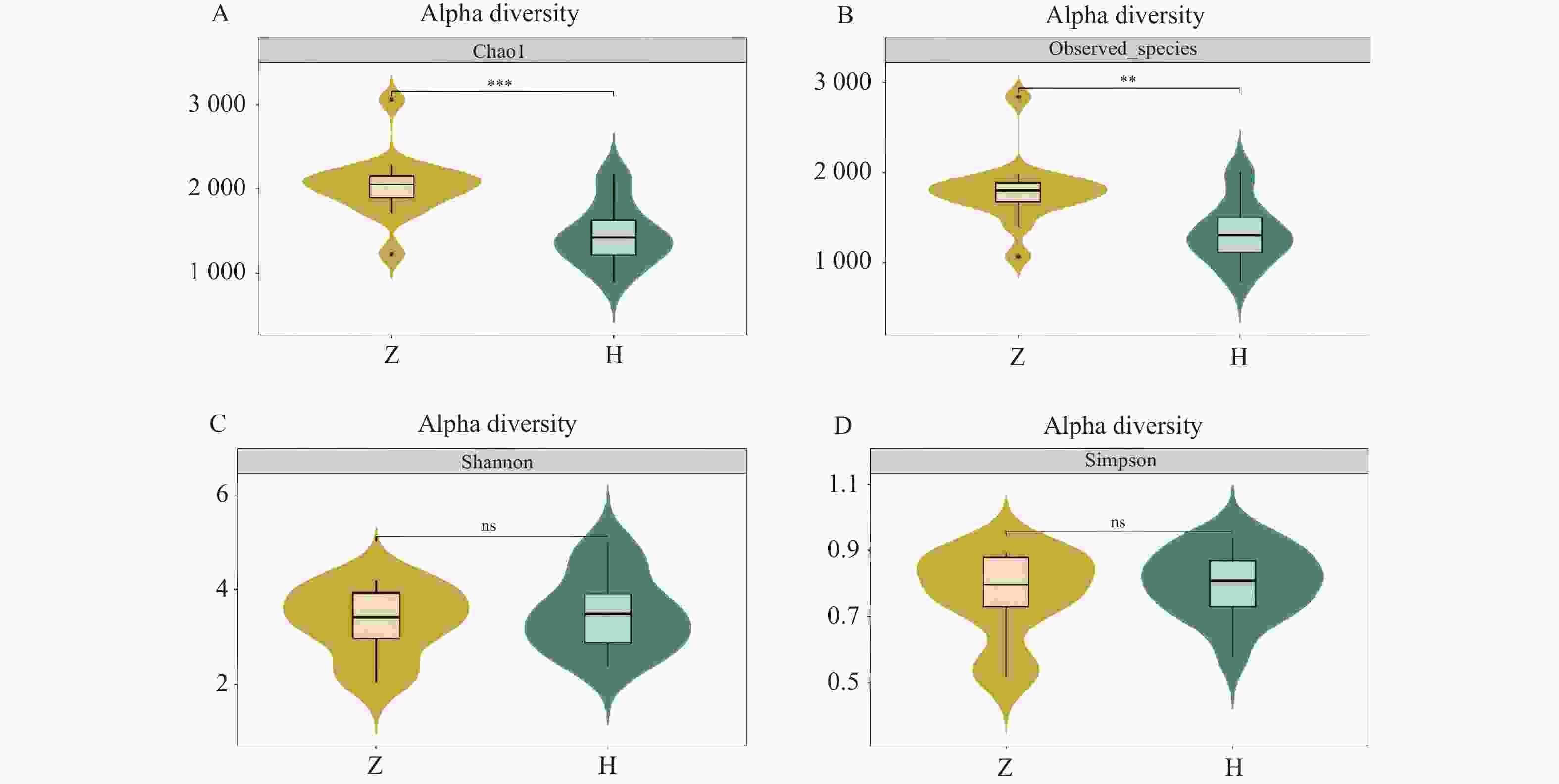
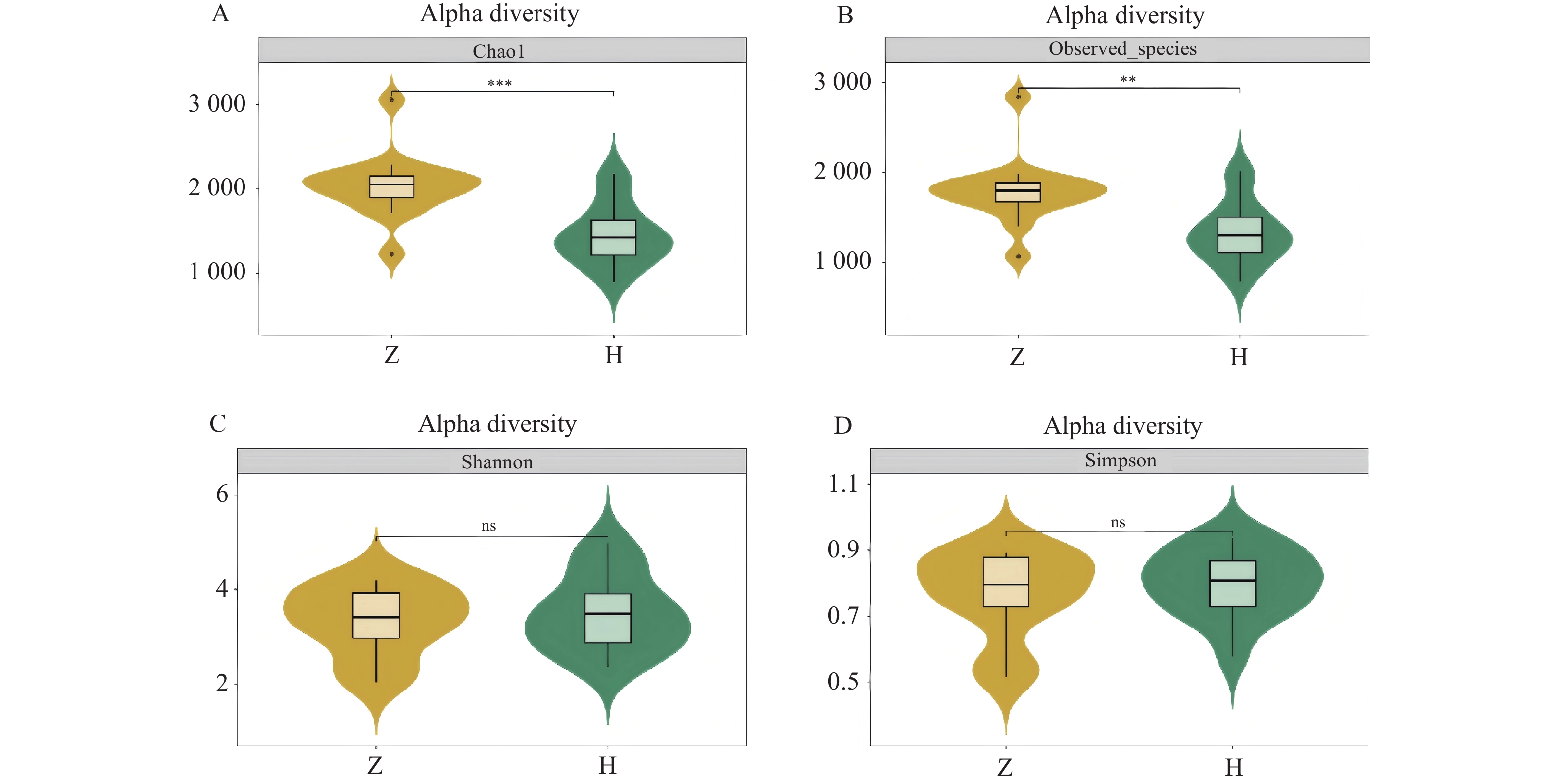
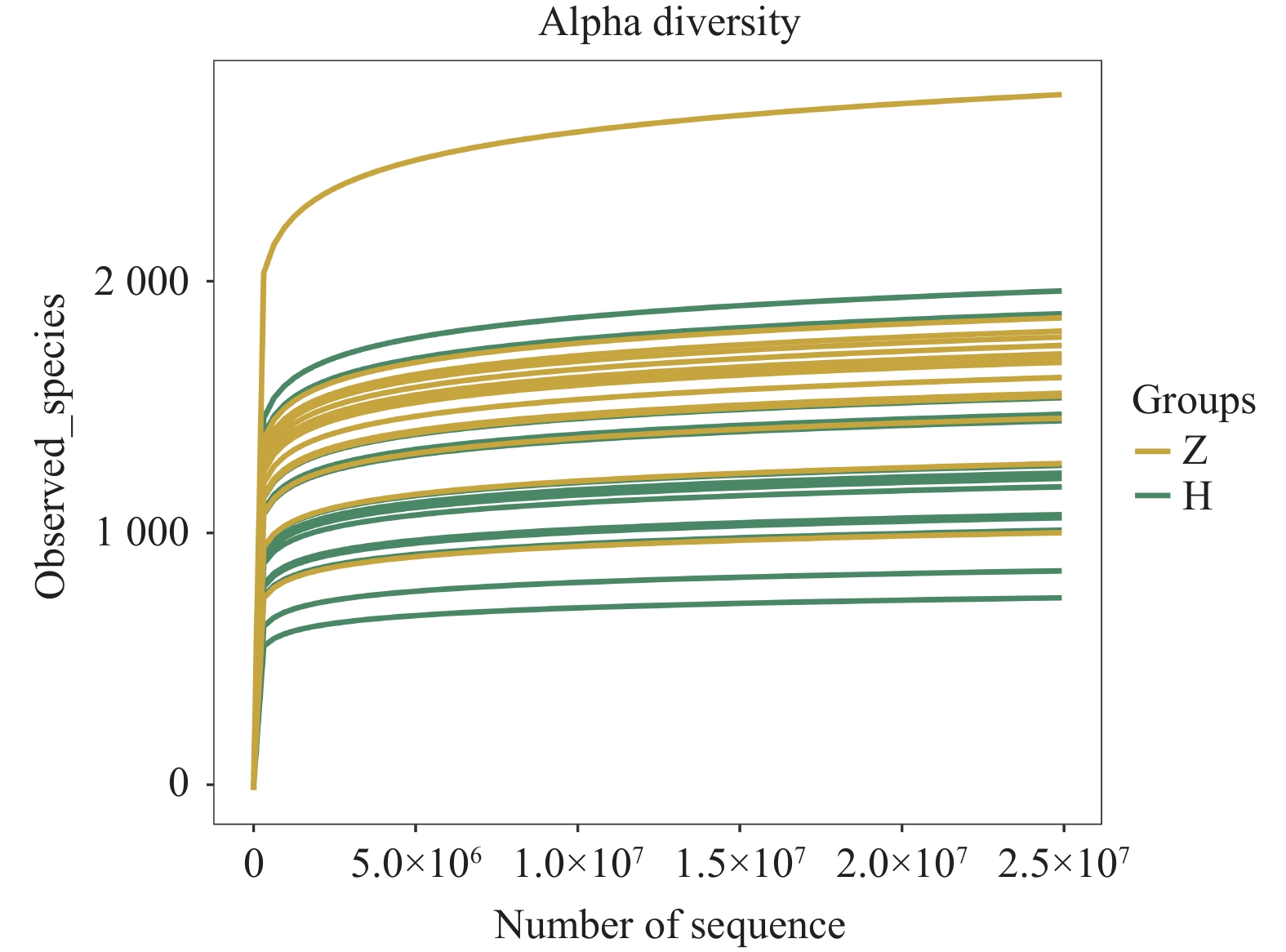
图 3 组间Alpha多样性小提琴图
A:chao 1指数;B:observed species 指数;C:shannon指数;D:simpson指数。
Figure 3. Violin plot of Alpha diversity between groups
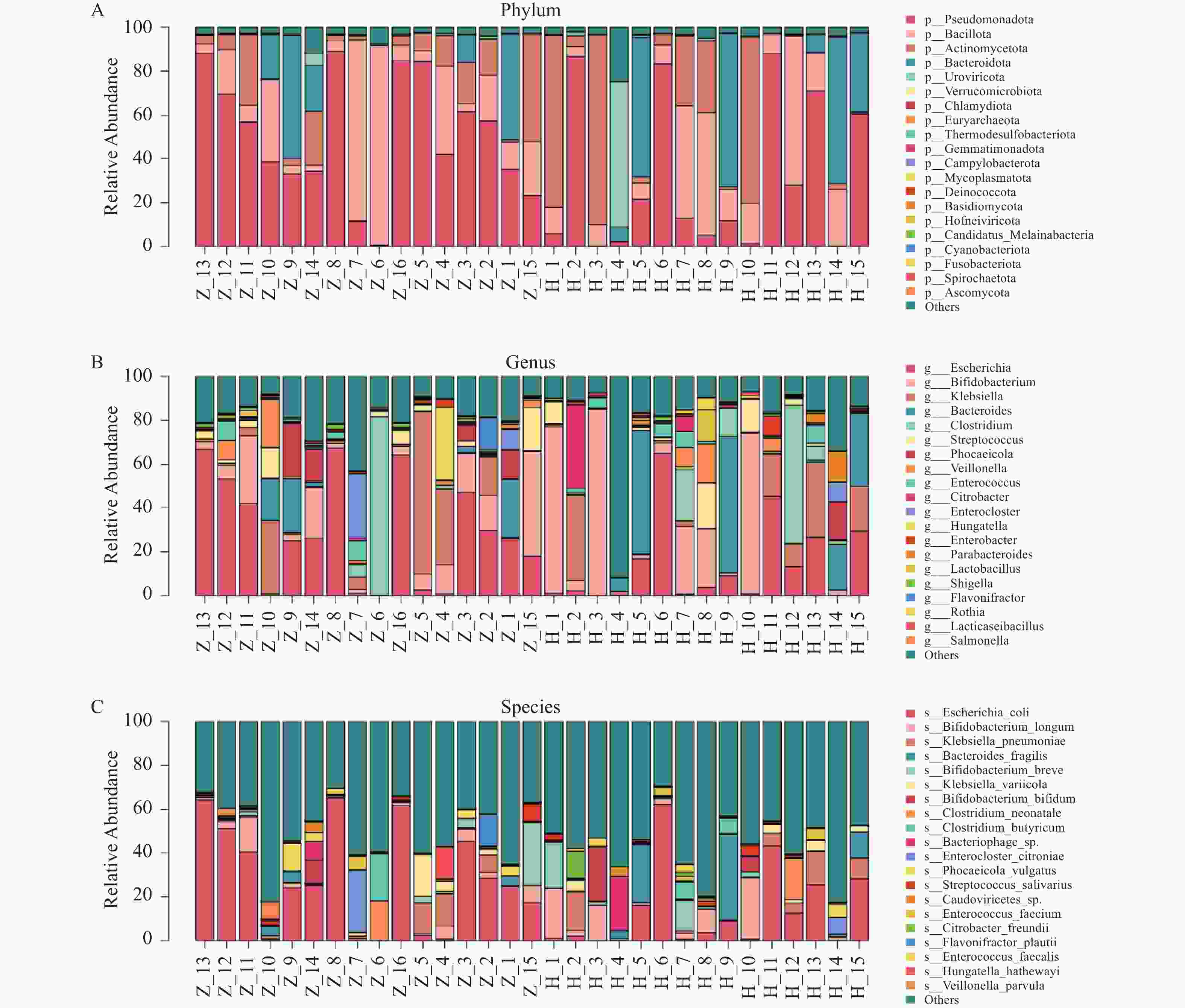
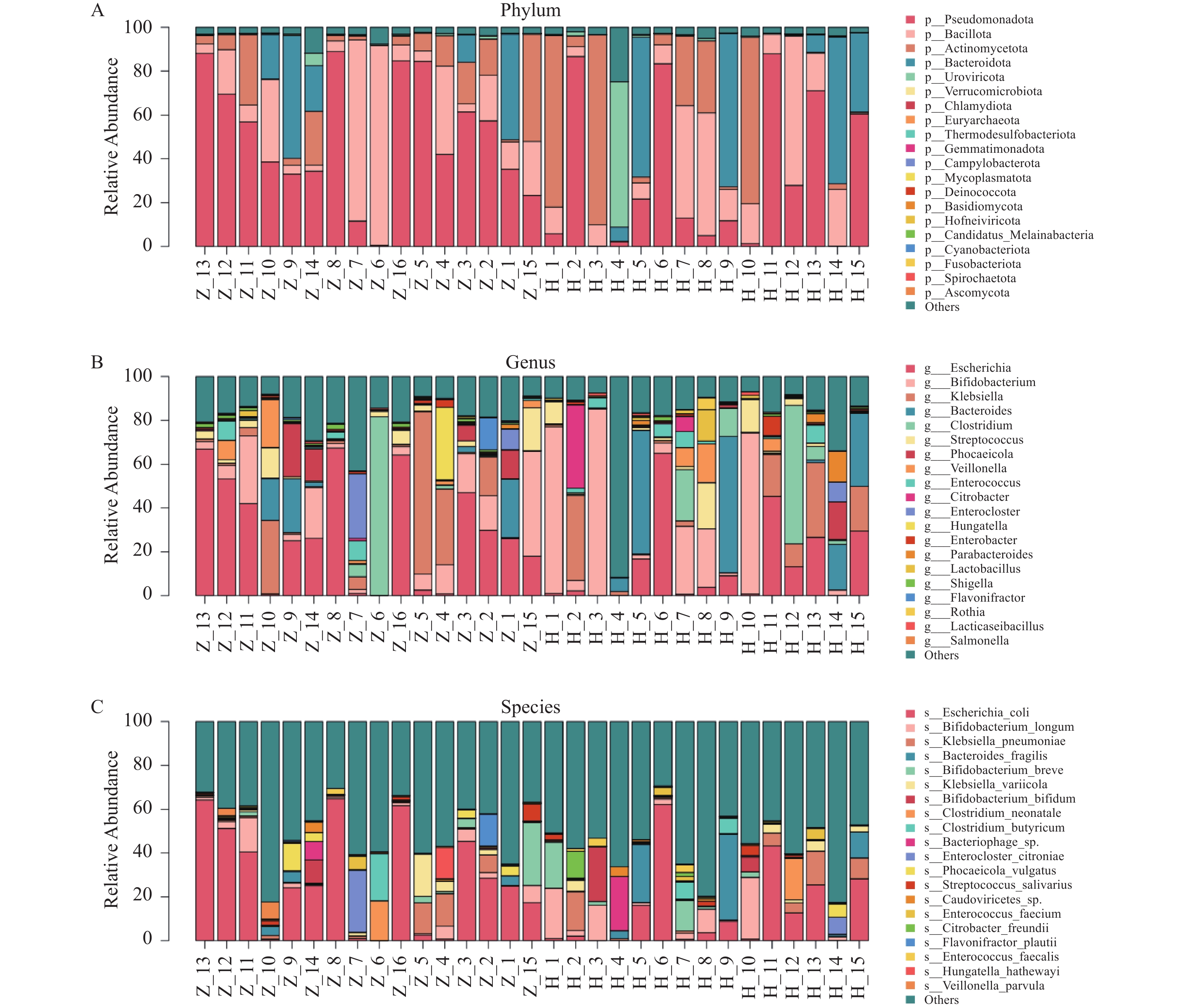
图 5 全样本菌落柱状堆叠图
A:全样本门层级菌落结构柱状图;B:全样本属层级菌落结构柱状图;C:全样本种层级菌落结构柱状图
Figure 5. Column stacking diagram in the full sample
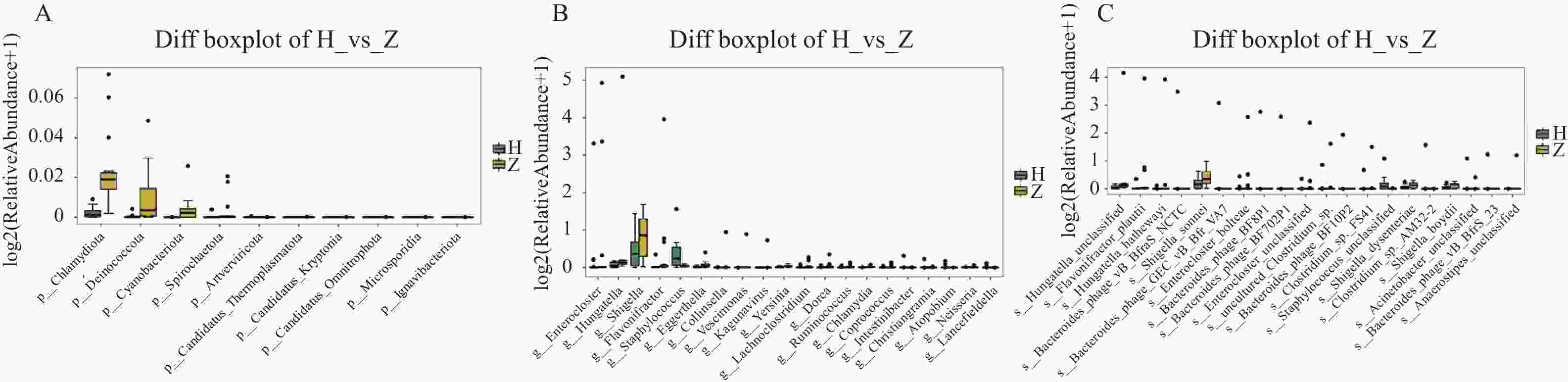
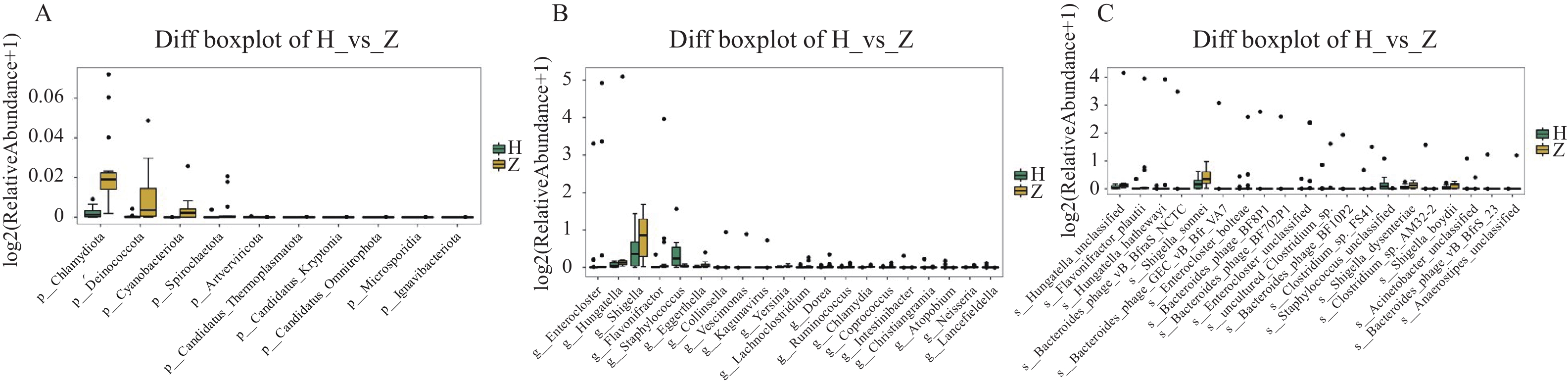
图 6 组间显著差异物种箱线图
A:门水平组间显著差异物种箱线图;B:属水平组间显著差异物种箱线图;C:种水平组间显著差异物种箱线图。
Figure 6. Boxplots of significantly different species between groups
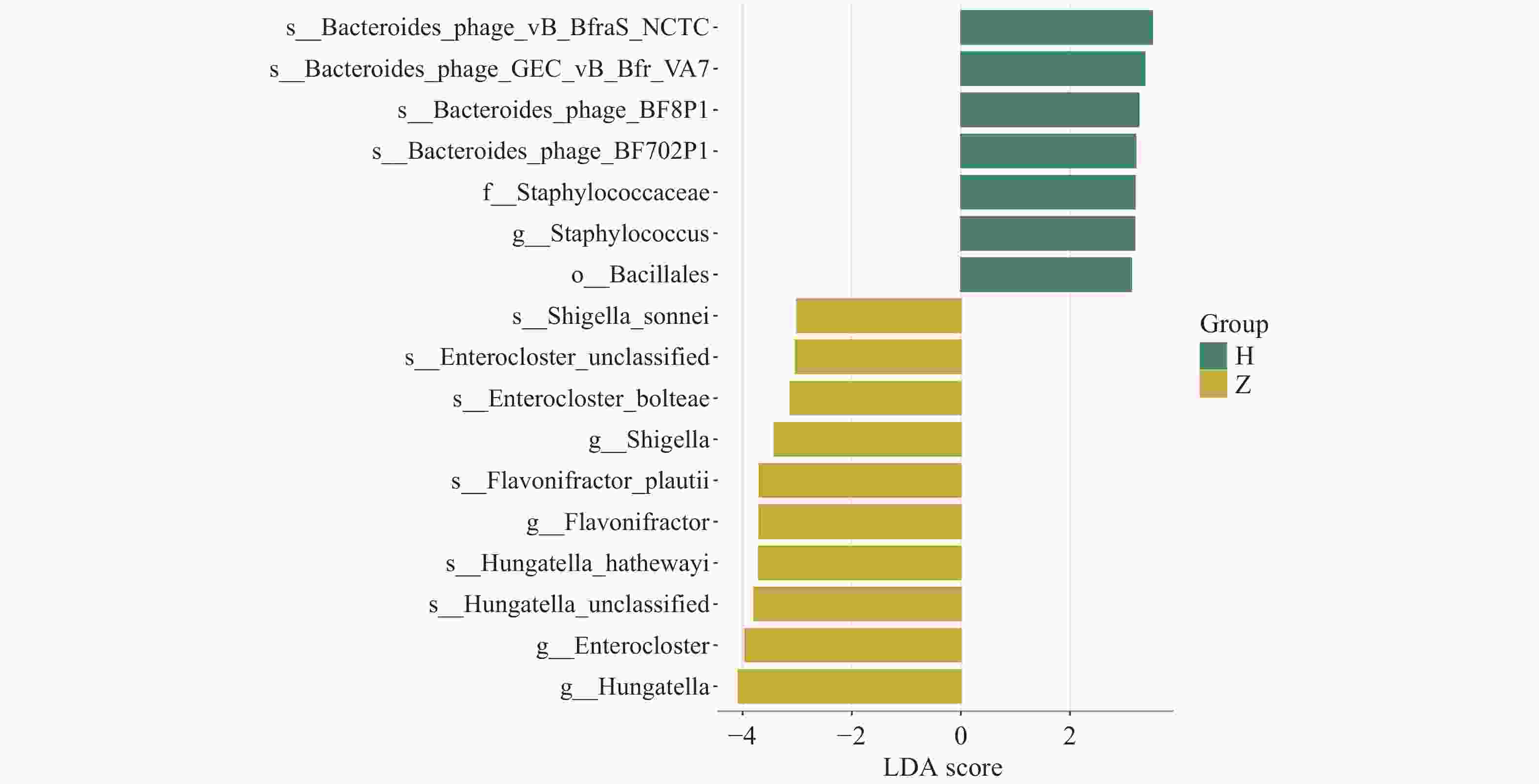
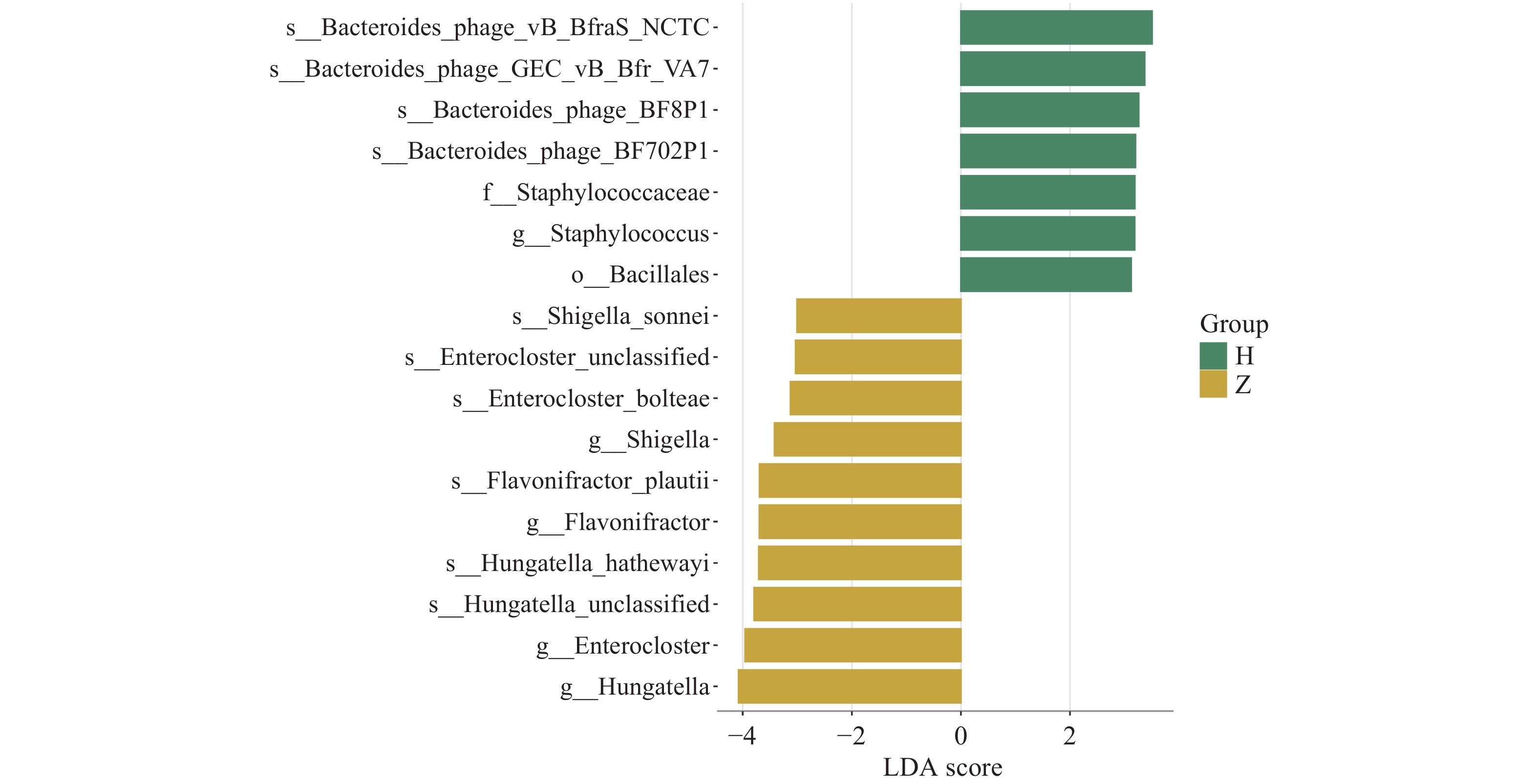
图 7 LDA分布柱状图
注:图中柱子的高低反映LDA值的大小,柱子越高,LDA值越大,该物种组间的差异贡献度越大,不同的颜色代表该物种所富集的分组。
Figure 7. Histogram of LDA distribution
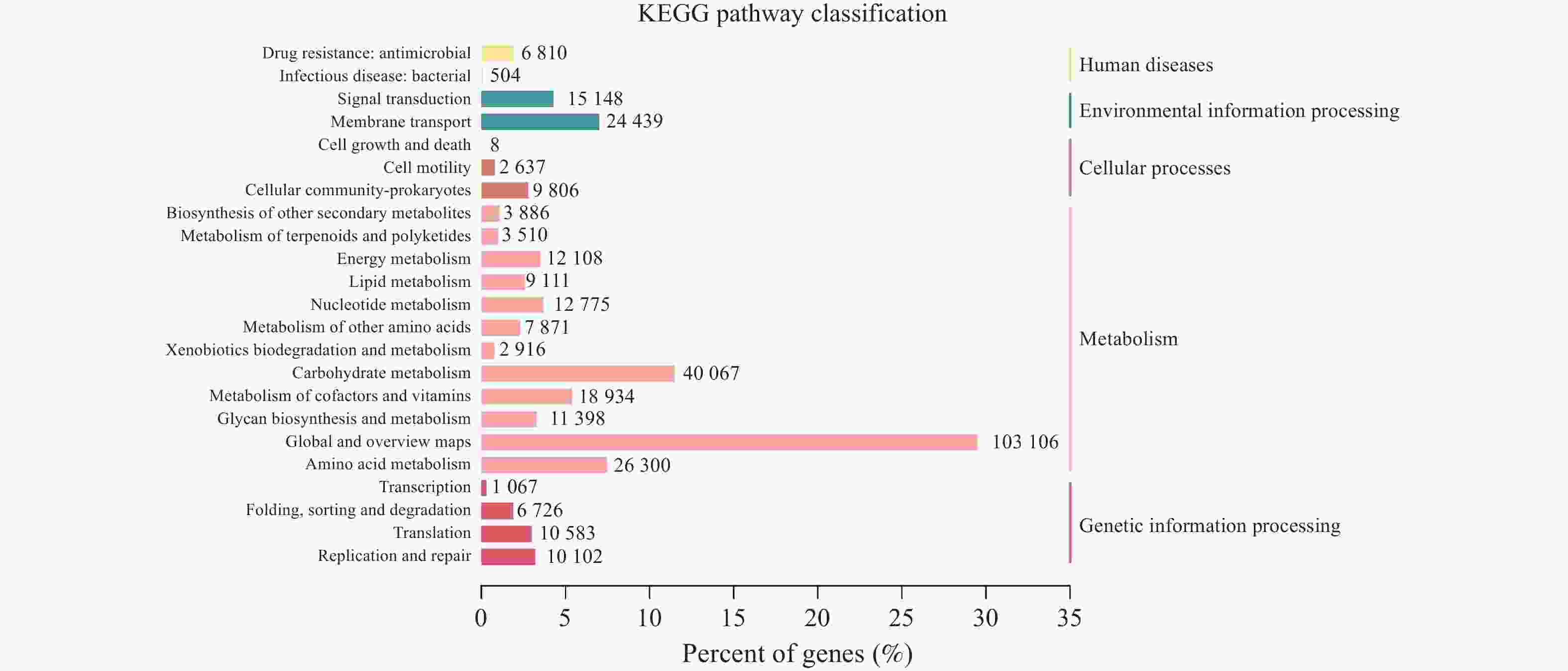
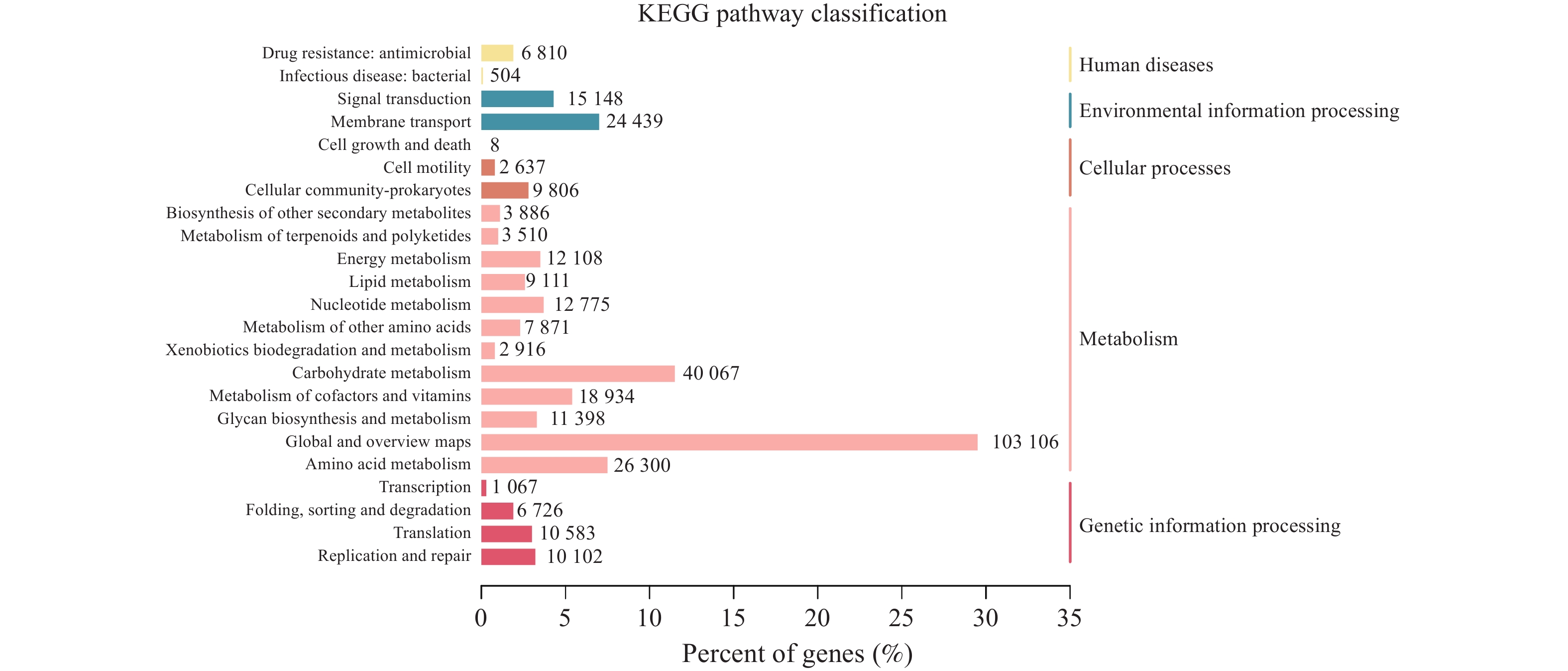
图 10 KEGG通路注释结果统计图
注:左边纵坐标为KEGG PATHWAY的二级分类信息,右边纵坐标为一级分类信息,横坐标代表注释到各二级分类信息的Unigene数量所占的百分比,柱子上的数字表示注释到各二级分类信息的Unigene数目。
Figure 10. KEGG Statistical plot of pathway annotation results
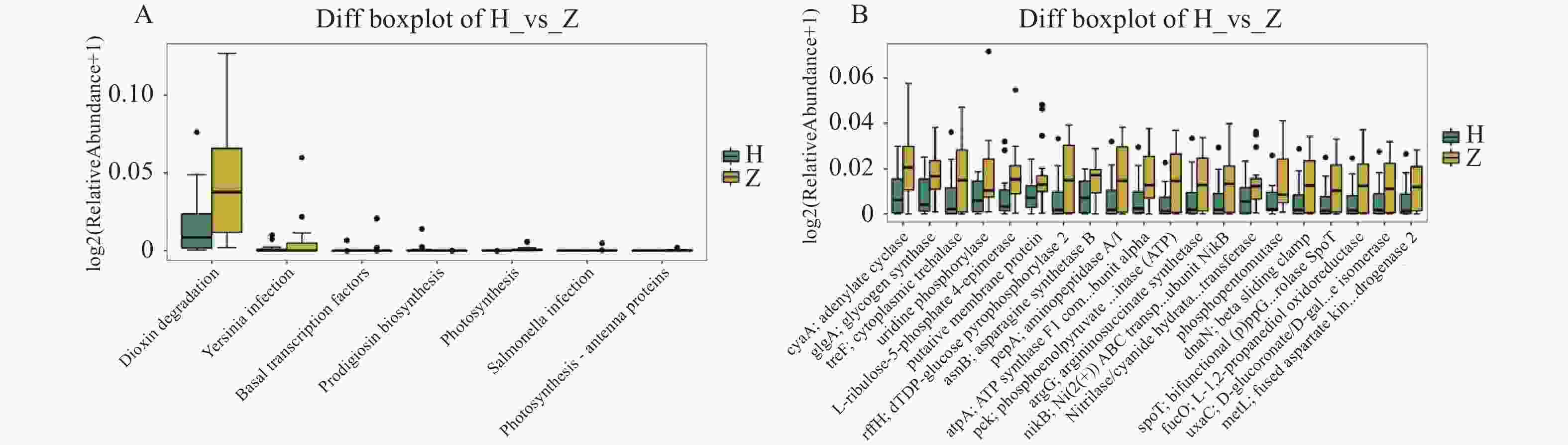
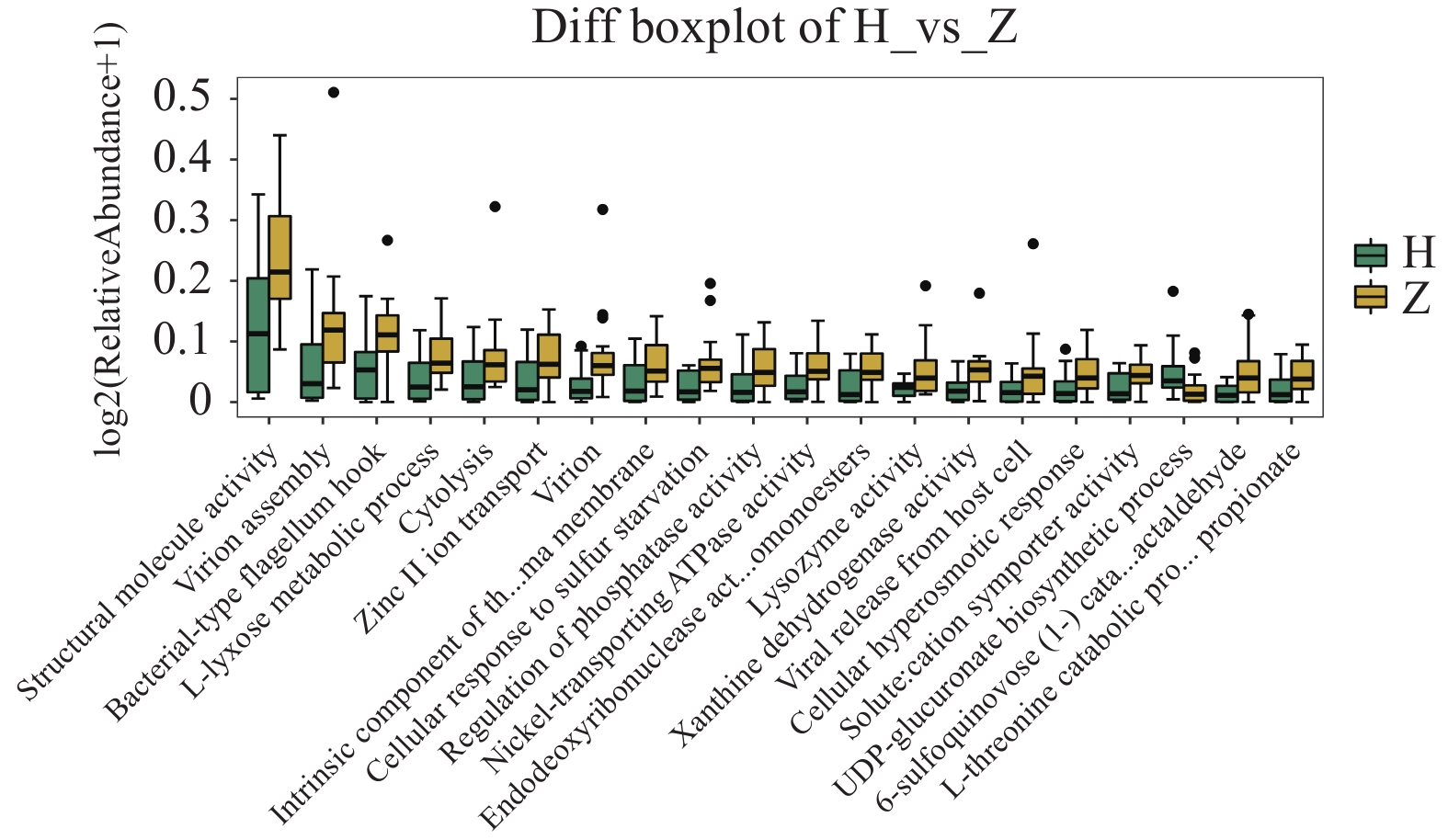
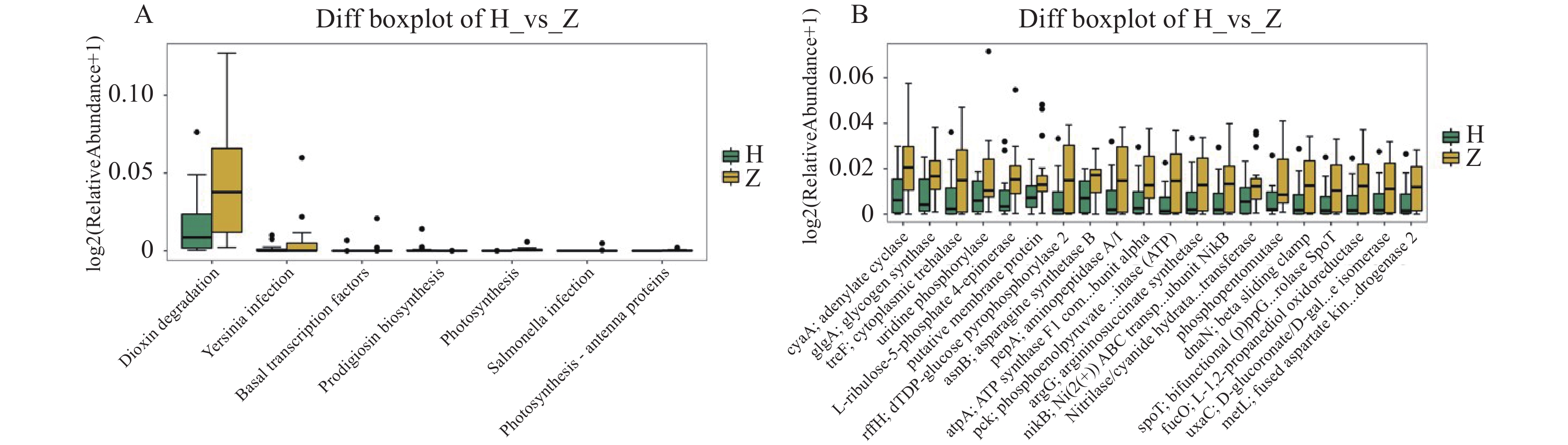
图 11 组间差异代谢功能箱线图
A:第3水平组间差异代谢功能箱线图;B:第4水平组间差异代谢功能箱线图。
Figure 11. Boxplot of differential metabolic function between groups
表 2 研究对象的基本特征表[($ \bar x \pm s $)/ n(%) / M(P25,P75)]
Table 2. Basic characteristics of the study subjects[($ \bar x \pm s $)/ n(%) / M(P25,P75)]
资料 Z组(n = 16) H组(n = 15) t / χ2 / Z P 母亲 年龄(岁) 31.9 ± 4.4 30.0 ± 3.1 1.362 0.184 孕前BMI(kg/m2) 20.6 ± 1.8 21.8 ± 3.6 −1.229 0.233 孕期体重增加数(kg) 12.5 ± 4.6 14.5 ± 4.8 −1.183 0.246 胎龄 273.6 ± 8.4 270.9 ± 8.4 0.897 0.377 民族(汉族/少数民族) 14/2 14/1 0.301 0.583 饮食习惯 肉类、奶类、蛋类、豆制品、蔬菜、米面等均食用 9(56.25) 9(60) 0.045 0.833 未进食其中一类及以上 7(43.75) 6(40) 睡眠时间 3~4 h 0(0) 1(6.67) 1.393 0.707 5~6 h 6(37.5) 5(33.33) 7~9 h 8(50) 8(53.33) 10 h及以上 2(12.5) 1(6.67) 情绪状态 愉快 9(65.25) 8(53.33) 0.338 0.844 一般 5(31.25) 4(26.67) 焦虑 2(12.5) 3(20) 产前工作情况 工作至产前 10(62.5) 11(73.33) 0.683 0.877 一直未工作 2(12.5) 1(6.67) 孕早期未工作 2(12,5) 1(6.67) 孕晚期未工作 2(12.5) 2(13.33) 家庭月收入(元) 3000 ~5000 3(18.75) 4(26.67) 0.313 0.855 5000 ~8000 4(25) 3(20) > 8000 9(56.25) 8(53.33) 哺乳频次 2~3 h 10(62.5) 9(60) 0.02 0.99 3~4 h 5(31.25) 5(33.33) 4~5 h 1(6.25) 1(6.67) 母乳低聚糖含量 二糖(μg/mL) 21.76 ± 13.43 19.30 ± 10.21 0.572 0.572 三糖(μg/mL) 1.28 ± 0.52 1.66 ± 0.79 −1.588 0.125 四糖(μg/mL) 0.70(0.58,5.19) 0.59(0.41,5.77) −0.395 0.693 婴儿 性别(男/女) 9/7 6/9 0.819 0.366 出生方式(顺/剖) 9/7 13/2 − 0.142 日龄(d) 42.3 ± 9.4 40.3 ± 9.7 0.595 0.556 出生体重(g) 3047.9 ± 355.53155.7 ± 407.1−0.787 0.438  下载: 导出CSV
下载: 导出CSV
表 1 生信统计学方法
Table 1. Bioinformatic methods
分析步骤 具体方法/R包 适用条件 差异阈值 Alpha多样性分析 Vegan包(R 3.6.0) 计算Chao1、Shannon、Simpson、Goods coverage指数,评估组内物种丰富度与均匀度 P < 0.05(Kruskal-Wallis检验组间差异) Beta多样性分析 Vegan包(R 3.6.0) 基于Bray-Curtis距离矩阵,通过PCoA可视化组间微生物群落结构差异 P < 0.05(Adonis置换检验) 差异物种分析 Wilcoxon rank-sum test / Kruskal-Wallis test 具备生物学重复的两组或多组独立样本整体差异检验 P < 0.05且|log2(fold_change)|>1 生物标志物筛选 LEfSe分析 筛选分类层级(门-种)的组间差异物种,结合LDA效应值与进化分支图可视化 LDA>3.0且P < 0.05 物种相关性网络 Ggnetwork+ggplot2(R 3.6.0) 种水平物种Spearman相关性分析,构建共发生网络并可视化节点与边的显著性 P < 0.05(相关性系数|r|>0.6) P值均采用Benjamini-Hochberg方法进行了False Discovery Rate校正,设定FDR阈值为0.05,当P < 0.05且q < 0.05时认为有统计学意义。
下载: 导出CSV
表 3 有效数据统计表
Table 3. Statistics of valid data
样本名称 原始数据 有效数据 有效比值
(百分比)Q20% Z_13 60250162 56705654 94.12 97.55 Z_12 37196126 35618524 95.76 97.67 Z_11 55343948 52321882 94.54 97.53 Z_10 49447642 47221864 95.50 97.79 Z_9 45170008 43274156 95.80 97.78 Z_14 39837914 38505026 96.65 98.17 Z_8 45344334 43768196 96.52 97.99 Z_7 46360094 44301642 95.56 98.12 Z_6 37217046 35013674 94.08 97.57 Z_16 32265088 31583084 97.89 97.89 Z_5 50265802 47508938 94.52 98.01 Z_4 52714104 50399352 95.61 97.88 Z_3 53897968 51168988 94.94 97.58 Z_2 60248614 57561724 95.54 97.73 Z_1 39366206 37976196 96.47 98.24 Z_15 39268806 35979202 91.62 97.97 H_1 42414996 41478490 97.79 98.39 H_2 54777350 54777350 97.82 98.33 H_3 37881744 36373614 96.02 98.23 H_4 60303778 59374764 98.46 98.61 H_5 60323554 59245992 98.21 98.61 H_6 45773860 44780848 97.83 98.50 H_7 43802314 42777414 97.66 98.61 H_8 40876800 40003914 97.86 98.62 H_9 60519990 59525040 98.36 98.77 H_10 48885328 47748062 97.67 98.38 H_11 51600084 50384310 97.64 98.59 H_12 60377958 59340358 98.28 98.68 H_13 51612786 50563974 97.97 98.55 H_14 60048128 59101428 98.42 98.99 H_15 50303208 49276774 97.96 98.48
下载: 导出CSV
-
[1] Cui M, Guo Q, Zhao S, et al. An untargeted comparative metabolomics analysis of infants with and without late-onset breast milk jaundice[J]. PLoS One, 2024, 19(8): e0308710. [2] Gao C, Guo Y, Huang M, et al. Breast milk constituents and the development of breast milk jaundice in neonates: A systematic review[J]. Nutrients, 2023, 15(10): 2261. [3] Par E J, Hughes C A, DeRico P. Neonatal hyperbilirubinemia: Evaluation and treatment[J]. Am Fam Physician, 2023, 107(5): 525-534. [4] Huang H, Huang J, Huang W, et al. Breast milk jaundice affects breastfeeding: From the perspective of intestinal flora and SCFAs-GPR41/43[J]. Front Nutr, 2023, 10: 1121213. doi: 10.3389/fnut.2023.1121213 [5] Wiciński M, Sawicka E, Gębalski J, et al. Human milk oligosaccharides: Health benefits, potential applications in infant formulas, and pharmacology[J]. Nutrients, 2020, 12(1): 266. [6] Xu D, Wan F, University J H, et al. Breastfeeding and infant gut microbiota: Influence of bioactive components[J]. Gut Microbes, 2025, 17: 2446403. [7] Su H, Yang S, Chen S, et al. What happens in the gut during the formation of neonatal jaundice—Underhand manipulation of gut microbiota?[J]. Int J Mol Sci, 2024, 25(16): 8582. [8] 熊晶晶, 张园园, 李檬, 等. 迟发型母乳性黄疸与婴儿排便频次的相关性[J]. 昆明医科大学学报, 2022, 43(9): 34-37. [9] 陈艳, 李檬, 张园园, 等. 纯母乳喂养儿迟发型母乳性黄疸与乳汁中3种功能性低聚糖水平及婴儿排便频次的相关性分析[J]. 中国妇幼保健, 2023, 38(3): 430-435. [10] Guo Q, Liu X, Cui M, et al. Characteristics of intestinal microbiota in infants with late-onset breast milk jaundice[J]. Front Nutr, 2023, 10: 1119768. [11] 贾尔肯阿依·阿不都拉依, 寇新红, 陈凤娟, 等. 迟发型母乳性黄疸患儿肠道菌群特征研究[J]. 中国妇幼卫生杂志, 2022, 13(1): 57-61. [12] 李亚璇, 莫茜, 孙建华, 等. 母乳喂养新生儿早期重度高胆红素血症肠道菌群特征研究[J]. 临床儿科杂志, 2019, 37(5): 351-355. [13] 殷丽丽, 杨志伟. 新生儿高胆红素血症与肠道菌群稳态及相关影响因素分析[C]//第二届全国康复与临床药学学术交流会议论文集. 南京, 2022: 649-653. [14] Li Y, Shen N, Li J, et al. Changes in intestinal flora and metabolites in neonates with breast milk jaundice[J]. Front Pediatr, 2020, 8: 177. [15] Leung J W, Liu Y L, Leung P S C, et al. Expression of bacterial β-glucuronidase in human bile: An in vitro study[J]. Gastrointest Endosc, 2001, 54(3): 346-350. [16] Talukdar P K, Sarker M R. Characterization of putative sporulation and germination genes in Clostridium perfringens food-poisoning strain SM101[J]. Microorganisms, 2022, 10(8): 1481. [17] Yang X T, Wang J, Jiang Y H, et al. Insight into the mechanism of gallstone disease by proteomic and metaproteomic characterization of human bile[J]. Front Microbiol, 2023, 14: 1276951. [18] Harrison M A, Faulds-Pain A, Kaur H, et al. Clostridioides difficile para-cresol production is induced by the precursorpara-hydroxyphenylacetate[J]. J Bacteriol, 2020, 202(18): 1-16. [19] Zhang X, Zeng S, Cheng G, et al. Clinical manifestations of neonatal hyperbilirubinemia are related to alterations in the gut microbiota[J]. Children, 2022, 9(5): 764. [20] Zhou W, Cui Y, Chen M, et al. Production of bilirubin via whole-cell transformation utilizing recombinant Corynebacterium glutamicum expressing a β-glucuronidase from Staphylococcus sp. RLH1[J]. Biotechnol Lett, 2024, 46(2): 223-233. [21] 齐玉敏, 王友军. 新生儿胆汁淤积性黄疸血清胆红素水平与肠道菌群失调的相关性[J]. 临床荟萃, 2022, 37(12): 1117-1121. [22] Kim Y H, Lee D H, Seo H S, et al. Genome-based taxonomic identification and safety assessment of an Enterococcus strain isolated from a homemade dairy product[J]. Int Microbiol, 2024, 27(5): 1513-1525. [23] Kubašová I, Lauková A, Hamarová Ľ, et al. Evaluation of enterococci for potential probiotic utilization in dogs[J]. Folia Microbiol, 2019, 64(2): 177-187. [24] Duan M, Han Z H, Huang T, et al. Characterization of gut microbiota and short-chain fatty acid in breastfed infants with or without breast milk jaundice[J]. Lett Appl Microbiol, 2021, 72(1): 60-67. [25] Yan W, Du N, Zhang K, et al. Bilirubin-microbiota interaction: Molecular mechanisms and therapeutic strategies in neonatal jaundice[J]. Front Microbiol, 2026, 16: 1749152. [26] Wu Q, Guo Q, Yang F, et al. Heterologous expression and enzymatic properties of β-glucuronidase from Clostridium perfringens and its application in bilirubin transformation[J]. Microorganisms, 2025, 13(5): 1043. [27] Jain V, Dalby M J, Alexander E C, et al. Association of gut microbiota and gut metabolites and adverse outcomes in biliary atresia: A longitudinal prospective study[J]. Hepatol Commun, 2024, 8(11): e0550. [28] Bai Y, Zhao T, Gao M, et al. A novel gene alignment in Dorea sp. AM58-8 produces 7-dehydroxy-3β bile acids from primary bile acids[J]. Biochemistry, 2022, 61(24): 2870-2878. [29] Sinha S R, Haileselassie Y, Nguyen L P, et al. Dysbiosis-induced secondary bile acid deficiency promotes intestinal inflammation[J]. Cell Host Microbe, 2020, 27(4): 659-670. e5. [30] Jin M, Cui J, Ning H, et al. Alterations in gut microbiota and metabolite profiles in patients with infantile cholestasis[J]. BMC Microbiol, 2023, 23(1): 357. [31] Li M, Li K, Tang S, et al. Restoration of the gut microbiota is associated with a decreased risk of hepatic encephalopathy after TIPS[J]. JHEP Rep, 2022, 4(5): 100448. [32] You J J, Qiu J, Li G N, et al. The relationship between gut microbiota and neonatal pathologic jaundice: A pilot case-control study[J]. Front Microbiol, 2023, 14: 1122172. [33] Shen J, Zhang J, Mo L, et al. Large-scale phage cultivation for commensal human gut bacteria[J]. Cell Host Microbe, 2023, 31(4): 665-677. e7. [34] Campbell D E, Ly L K, Ridlon J M, et al. Infection with Bacteroides Phage BV01 alters the host transcriptome and bile acid metabolism in a common human gut microbe[J]. Cell Rep, 2020, 32(11): 108142. [35] Campbell D E, Ly L K, Ridlon J M, et al. Infection with Bacteroides Phage BV01 alters the host transcriptome and bile acid metabolism in a common human gut microbe[J]. Cell Rep, 2020, 32(11): 108142. [36] Han W, Huang C, Zhang Q, et al. Alterations in gut microbiota and elevated serum bilirubin in primary biliary cholangitis patients treated with ursodeoxycholic acid[J]. Eur J Clin Investig, 2022, 52(2): e13714. [37] Ratliff M, Zhu W, Deshmukh R, et al. Homologues of neisserial heme oxygenase in gram-negative bacteria: Degradation of heme by the product of thepigAGene ofPseudomonas aeruginosa[J]. J Bacteriol, 2001, 183(21): 6394-6403. [38] Wilks A, Ikeda-Saito M. Heme utilization by pathogenic bacteria: Not all pathways lead to biliverdin[J]. Acc Chem Res, 2014, 47(8): 2291-2298. [39] Becker P, Naughton F, Brotherton D, et al. Mechanism of substrate binding and transport in BASS transporters[J]. eLife, 2023, 12: RP89167. [40] Li Z, Zhang Y, Luo X, et al. Dynamic relationships between bilirubin concentrations and the gut microbiota in the neonatal period: A pilot prospective cohort study[J]. Pediatr Investig, 2025, 9(4): 347-360. [41] 熊佳, 曾加, 周小仙, 等. 益生菌辅助治疗代谢功能障碍相关脂肪性肝病的研究进展[J]. 昆明医科大学学报, 2025, 46(7): 163-174. -


 点击查看大图
点击查看大图
计量
- 文章访问数: 10
- HTML全文浏览量: 10
- PDF下载量: 2
- 被引次数: 0
